


Abstract
The γ-aminobutyric acid (GABA)A receptor is a hetero-oligomer consisting of five subunits, the combination of which confers unique pharmacological properties to the receptor. To understand the physiological role of native GABAAreceptors, it is critical to determine their subunit compositions. The pharmacological characteristics of human α5β3γ2 and α5β3γ3 GABAA receptors stably expressed in L(tk−) cells were characterized with the α5-selective ligand [3H]L-655,708 and compared with the pharmacological characteristics of [3H]L-655,708 binding sites from rat and human hippocampus. Saturation analyses revealed a 9-fold selective affinity of [3H]L-655,708 for α5β3γ2 receptors (K d = 1.7 ± 0.4 nm), compared with α5β3γ3 receptors (K d = 15 ± 3 nm). Rat and human hippocampal [3H]L-655,708 binding sites had affinities of 2.2 ± 0.6 and 1.0 ± 0.2 nm, respectively, comparable to the affinity of α5β3γ2 receptors. Pharmacological analysis of [3H]L-655,708 binding sites in rat and human hippocampi revealed a strong correlation with the affinities of seven benzodiazepine site ligands for α5β3γ2 but not α5β3γ3 receptors. Immunoprecipitation of [3H]L-655,708 binding sites from rat hippocampus with a γ2-selective antibody yielded 19 ± 4% of total benzodiazepine binding sites measured using [3H]Ro15–1788, whereas no specific binding was measured after immunoprecipitation with an anti-γ3 antibody. Combinatorial immunoprecipitations of [3H]muscimol binding sites with anti-α5 and anti-γ2 or anti-α5 and anti-γ3 antibodies established the preferential expression of α5γ2 receptors, accounting for 22 ± 2% of total rat hippocampal GABAA receptors. These observations provide pharmacological and structural evidence for the prevalence of α5β3γ2 GABAA receptors in rat hippocampus, despite the clustering of α5 and γ3 loci on the same chromosome.
The GABAA receptor is the main inhibitory ligand-gated ion channel in the central nervous system. It contains modulatory sites for endogenous molecules such as the neurosteroids, as well as for many therapeutic drugs, such as barbiturates, anesthetics, and benzodiazepines (Sieghart, 1995). It is now generally accepted that the GABAA receptor is a pentameric protein with an integral chloride ion channel formed by the second transmembrane domain of each of the five subunits. A family of GABAA receptor subunits (α1–α6, β1–β3, γ1–γ3, δ, and ε) have been identified in mammalian brain using molecular cloning techniques (for review, see McKernan and Whiting, 1996; Davies et al., 1997; Whiting et al., 1997). At least one α subunit, one β subunit, and one γ subunit are required to form fully functional receptors in vivo(Pritchett et al., 1989), and the combination of α and γ subunits is a crucial determinant of the properties of the benzodiazepine binding site (Hadingham et al., 1993; Waffordet al., 1993; Luddens et al., 1994; Benkeet al., 1996). The α1 subunit-containing receptors exhibit BZ1-type pharmacological characteristics, characterized by a high affinity for zolpidem, whereas α2, α3, and α5 subunits are present in BZ2-type receptors, which have a low affinity for zolpidem (Pritchett et al., 1989; Pritchett and Seeburg, 1990). A third class of GABAA receptors also exists; these receptors contain an α4 or α6 subunit and have a low affinity for most of the classical benzodiazepines (Luddens et al., 1990;Wisden et al., 1991; Wafford et al., 1996; Benkeet al., 1997).
Receptors expressing an α5 subunit together with β and γ2 subunits in cell lines are distinguished from BZ1 receptors by their low affinity for zolpidem (Pritchett and Seeburg, 1990; Luddenset al., 1994) and from other BZ2 receptors by their 10–20-fold higher affinities for Ro15–4513 (Hadingham et al., 1993; Luddens et al., 1994) and for several 8-substituted benzodiazepines (Gillard et al., 1994). Thus, α5-containing receptors have a unique pharmacological profile.
In the rat central nervous system, α5 subunit-containing receptors have restricted and well defined expression. In situhybridization and immunocytochemistry studies have shown that this subtype is present in abundance in the CA1 and CA3 fields of the hippocampus, is present to a lesser extent in the cortex and olfactory bulb, and is virtually absent in other regions of the brain (Wisdenet al., 1992; Fritschy and Mohler, 1995; Quirk et al., 1996).
The exact subunit composition of receptors containing an α5 subunitin vivo is not known. The colocalization of α5, β3, and γ3 subunits on chromosome 15 suggests possible associations among these subunits, and deletion of this locus reduces zolpidem-insensitive, radiolabeled benzodiazepine binding (Nakatsuet al., 1993). On the other hand, the pharmacological characteristics of α5-containing receptors immunoprecipitated from rat brain are closer to those demonstrated in cells transfected with α5β3γ2 than those observed with α5β3γ3 (McKernan et al., 1991; Luddens et al., 1994). Similarly, electrophysiological studies indicated α5β3γ2L as the isoform expressed by hippocampal CA1 pyramidal cells (Burgard et al., 1996).
Using a 50–100-fold selective ligand for α5 subunit-containing receptors (Quirk et al., 1996), [3H]L-655,708, we describe here the pharmacological characteristics of α5 subunit-containing receptors in rat and human brain and compare them with those of stable cell lines expressing α5β3γ2 and α5β3γ3 receptors. Implications for the structure of native α5 subunit-containing receptors are discussed.
Experimental Procedures
Materials.
[3H]Ro15–1788 and [3H]muscimol were purchased from New England Nuclear-Du Pont (Hertfordshire, UK). [3H]L-655,708 (76.7 Ci/mmol) was prepared as previously described (Quirk et al., 1996). CL218872 and CGS8216 were gifts from Lederle and Ciba-Geigy/Novartis, respectively, and other benzodiazepine site ligands were from Sigma Biochemicals or Research Biologicals Inc.
Human tissues.
Hippocampi were from adult cadaveric brain tissue obtained from subjects without any neurodegenerative disease and without obvious morphological abnormalities of the hippocampi.
Membrane preparation and binding assays.
P2 membranes were prepared from brain regions of adult male rats or from cadaveric hippocampi from adult human subjects as previously described (McKernanet al., 1991). Radioligand binding assays were performed with brain membranes or membranes prepared from stably transfected cell lines (Hadingham et al., 1993; Sur et al., 1997), with [3H]L-655,708 (0.1–40 nm), in a final volume of 0.5 ml containing 50–100 μg of protein in 10 mm Tris·HCl, 1 mm EDTA, pH 7.4, at 4°. For saturation analyses with rat and human hippocampi, 1 μm zolpidem was added to the assays, to prevent binding to other α subunits at high [3H]L-655,708 concentrations. Similarly, immunoprecipitated receptor-protein A complexes were incubated with [3H]L-655,708 (20–24 nm), [3H]Ro15–1788 (20 nm), or [3H]muscimol (40 nm) for 1–2 hr at 4°. Nonspecific binding was defined with 10 μm flunitrazepam or 100 μm GABA, for 3H-benzodiazepine or [3H]muscimol binding, respectively. After 1–2-hr incubations at 4°, assay mixtures were filtered through Whatman GF/B filters using a cell harvester (Brandel) and were washed four times with cold buffer. Filters were immersed overnight in scintillation cocktail, and radioactivity was determined in a Beckman liquid scintillation counter. Data points were fitted by nonlinear regression analysis (Excel; Microsoft); for competition experiments, the Ki values were calculated according to the Cheng-Prusoff equation (Cheng and Prusoff, 1973).
Immunoprecipitation of GABAA receptors.
The antibodies used in this study were previously characterized and shown to be subunit specific (McKernan et al., 1991; Quirket al., 1994a, 1994b, 1995). Immunoprecipitation of receptors was carried out using antibodies to GABAA receptors, as previously described (McKernan et al., 1991; Quirk et al., 1994a). Briefly, 100 μl of protein A-Sepharose beads was incubated with 40–80 μl of antibody for 1 hr at room temperature. After washing with Tris-buffered saline (10 mm Tris·HCl, 150 mm NaCl) containing 0.1% Tween 20, beads were incubated overnight at 4° with deoxycholate (0.5%)-solubilized receptors from hippocampus. Beads were washed twice with Tris-buffered saline/Tween 20, and binding studies were performed using 25–50 μl of packed beads in each tube. Parallel experiments with an antibody directed against the 5-hydroxytryptamine3 receptor served as controls for the immunoprecipitation experiments.
Results
[3H]L-655,708 binding characteristics.
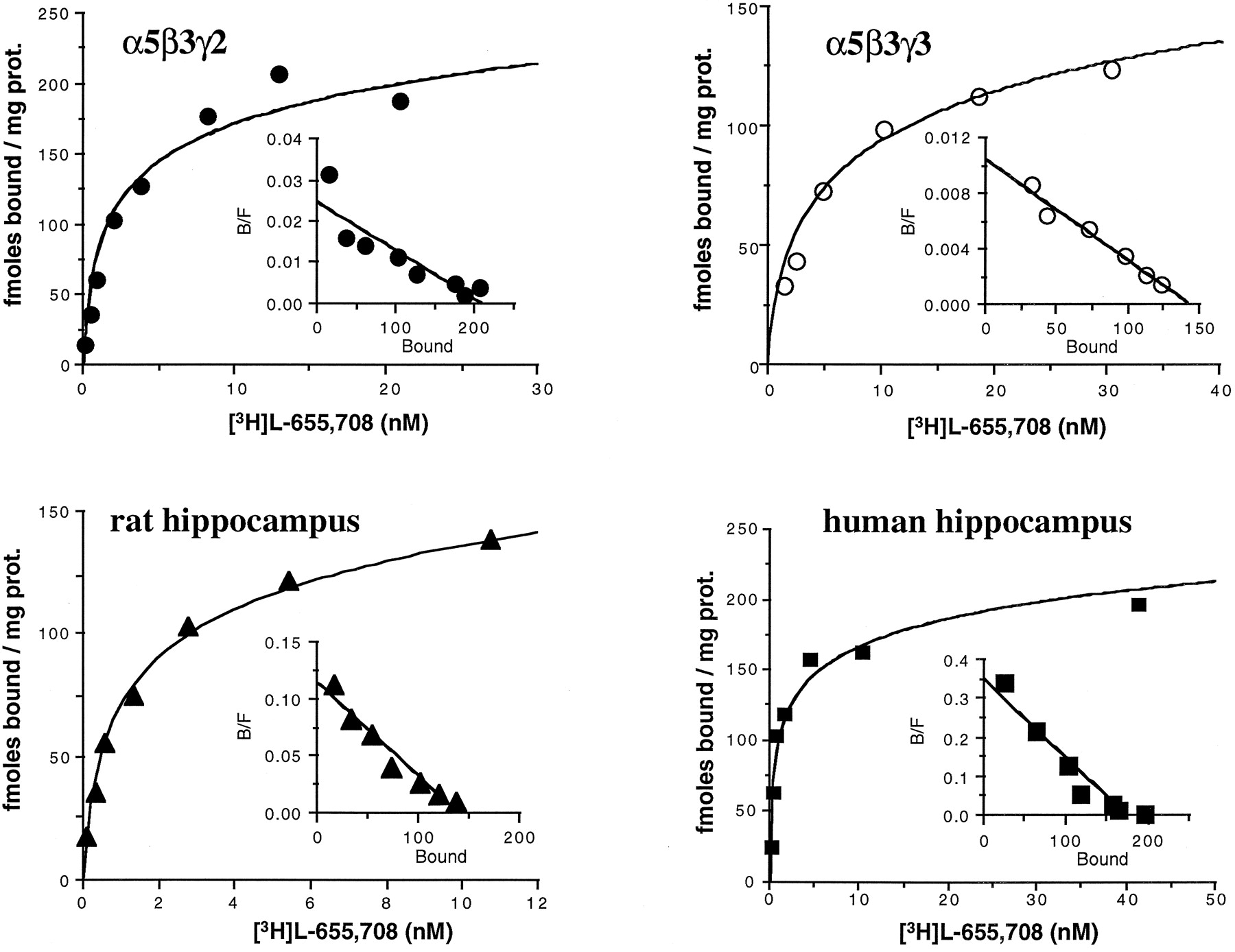
To determine the affinity of [3H]L-655,708, saturation experiments were performed with membranes prepared from α5β3γ2- or α5β3γ3-expressing cells, as well as rat and human hippocampi (Fig. 1 and Table1). They revealed a 9-fold selectivity of [3H]L-655,708 for α5β3γ2 versus α5β3γ3 receptors, withKd values of 1.7 ± 0.4 and 15 ± 3 nm, respectively. Saturation analysis of [3H]L-655,708 binding to rat and human hippocampi revealed the existence of a single high affinity binding site, with Kd values of 2.2 ± 0.6 and 1.1 ± 0.2 nm,respectively. These experiments also showed no difference (p > 0.46, t test) in the numbers of [3H]L-655,708 binding sites in human (340 ± 184 fmol/mg of protein, mean ± standard deviation, five experiments) and rat (251 ± 74 fmol/mg of protein, mean ± standard deviation, three experiments) hippocampi. Determination of the [3H]L-655,708 (2 nm)/[3H]Ro15–1788 (1.8 nm) ratio, however, indicated a slightly higher proportion of α5-containing receptors in human hippocampus, with a ratio of 0.28 ± 0.04 (mean ± standard error, two experiments), compared with a ratio of 0.15 ± 0.02 (mean ± standard error, six experiments) for rat hippocampus (p < 0.005, t test).

Saturable binding of [3H]L-655,708 to human recombinant α5β3γ2 and α5β3γ3 GABAA receptors and α5-containing receptors from rat and human hippocampal membranes. The linear Scatchard plots (insets) show that [3H]L-655,708 binds to a single population of sites. Data shown are from a representative experiment that was performed at least three times. The binding parameters for these experiments were as follows: α5β3γ2,K d = 2.4 nm,B max = 223 fmol/mg of protein; α5β3γ3,K d = 10.2 nm,B max = 186 fmol/mg of protein; rat hippocampal membranes, K d = 1.7 nm, B max = 170 fmol/mg of protein; human hippocampal membranes,K d = 0.92 nm,B max = 200 fmol/mg of protein.
Affinities of [3H]L-655,708 for α5-containing receptors
Evidence that rat and human α5-containing receptors display α5β3γ2 pharmacological characteristics.
Competition experiments using [3H]L-655,708 (2–3 nm) and seven representative benzodiazepine site ligands from different chemical series were carried out in cell lines and hippocampal membranes. The results (Table2) established the selectivity of some compounds for either α5β3γ2 (CGS8216, L-655,708, and diazepam) or α5β3γ3 (CL218872) receptors. More specifically, CGS8216 demonstrated 14-fold selectivity for α5β3γ2, whereas CL218872 exhibited 5-fold selectivity for α5β3γ3 receptors. Correlation plots demonstrated a good relationship between the affinities of these compounds for α5β3γ2 receptor-expressing cells and rat or human [3H]L-655,708 binding sites (Fig.2, A and B). Furthermore, there was an excellent correlation of the pharmacological characteristics of rat and human α5 receptors (Fig. 2C). In contrast, a nonsignificant correlation (p > 0.05, Spearman correlation) was observed for the pharmacological characteristics of hippocampal [3H]L-655,708 binding sites and the α5β3γ3 isoform (data not shown).
Affinities of benzodiazepine site ligands for α5-containing receptors

Logarithmic-logarithmic plots ofK i values (nm) (mean ± standard error) for seven benzodiazepine site ligands, showing the correlation (Spearman correlation) between the pharmacological characteristics of rat (A) (r 2 = 1) and human (B) (r 2 > 0.92) hippocampal α5-containing receptors and the α5β3γ2 isoform, as well as between rat and human hippocampal α5 receptors (C) (r 2 > 0.92). Numbers near symbols, compounds listed in Table2.
Pharmacological characteristics of immunoprecipitated rat hippocampal GABAA receptors.
As shown in Fig.3, solubilized and immunoprecipitated α5-containing GABAA receptors retained high affinity (Kd = 3.7 ± 1.3 nm) [3H]L-655,708 binding. To gain more insight into the structure of rat hippocampal α5-containing receptors, immunoprecipitation experiments were performed with several subunit-specific antibodies, and the binding of [3H]L-655,708 (20–24 nm) and [3H]Ro15–1788 (20 nm) was determined (Table3). Because of the high concentration of [3H]L-655,708 used, binding was determined in the presence of zolpidem (1 μm), to prevent [3H]L-655,708 binding with low affinity to α1, α2, and α3 subunits. Because we anticipated the lack of 3H-benzodiazepine binding to receptors precipitated with antibodies to γ1 or δ subunits (Ymeret al., 1990; Quirk et al., 1995), the binding of [3H]muscimol (which binds to the GABA binding site of GABAA receptors) was measured in parallel experiments, to confirm that immunoprecipitation had occurred. All benzodiazepine sites (95 ± 6%) immunoprecipitated with anti-α5 antibody, as determined with [3H]Ro15–1788, exhibited [3H]L-655,708 binding. Antibodies selective for α1, α2, or α3 subunits were able to precipitate 3, 6, and 7%, respectively, of [3H]Ro15–1788 binding sites that also bound [3H]L-655,708 (Table 3). These populations of receptors accounted for a small proportion of total GABAA receptors.
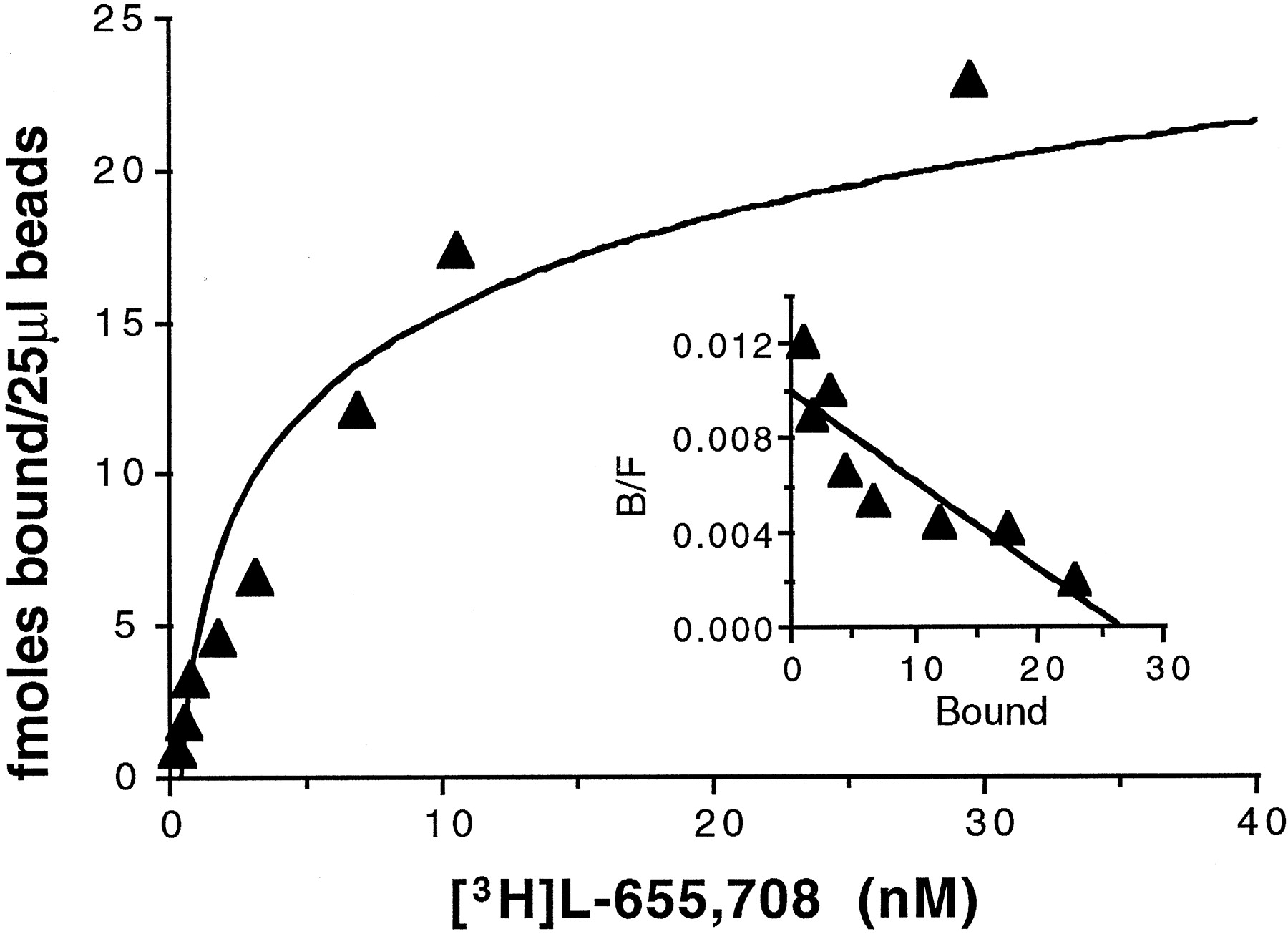
Saturation analysis of [3H]L-655,708 binding to anti-α5-immunoprecipitated receptors from rat hippocampus. The linear Scatchard plot (inset) indicates saturable binding of the radioligand to a single class of sites.
Percentage of [3H]L-655,708/[3H]Ro15-1788 binding in immunoprecipitated receptors from rat hippocampus
Among the three γ subunits assayed, only antibodies to γ2 immunoprecipitated [3H]L-655,708 binding sites. These α5γ2 subunit-containing receptors represented 19 ± 4% (mean ± standard error, five experiments) of total [3H]Ro15–1788 binding sites. The anti-γ3 antibody immunoprecipitated some [3H]Ro15–1788 binding sites (5.4 ± 2.4%, two experiments) but no [3H]L-655,708 binding, whereas no benzodiazepine binding was observed after precipitation with anti-γ1 or anti-δ antibodies.
Additional evidence for the presence of native α5β3γ2 receptors in rat hippocampus was provided by additive immunoprecipitation experiments with anti-α5, -γ2, and -γ3 antibodies, using [3H]muscimol binding (Fig.4). Solubilized GABAA receptors from hippocampus were immunoprecipitated either with a single antibody or with α5γ2 or α5γ3 combinations. As shown in Fig. 4, anti-γ2 antiserum immunoprecipitated the larger amount of [3H]muscimol binding sites (59%), whereas anti-α5 and anti-γ3 precipitated 31 and 7.6%, respectively. Interestingly, the antibody pair for α5γ2 immunoprecipitated 66% of [3H]muscimol binding sites, a proportion much smaller than the sum of anti-α5- and anti-γ2-precipitated sites (31 + 59 = 90%). In contrast, the quantity of [3H]muscimol binding sites immunoprecipitated by the pair for α5γ3 (38%) corresponds to the sum of individually precipitated receptors (31 + 7.6 = 38.6%). Results from another experiment yielded similar values for immunoprecipitated [3H]muscimol binding sites with antibodies for α5 (37%), γ2 (91%), γ3 (11%), α5γ2 (109%), and α5γ3 (49%). The difference between the calculated and measured values for the α5γ2 tandem indicates that this subunit combination accounts for 21.5 ± 2.5% (mean ± standard error, two experiments) of [3H]muscimol binding sites in rat hippocampus.

Immunoprecipitation of GABAA receptor binding sites from rat hippocampus by anti-α5, -γ2, and -γ3 antibodies. Binding of [3H]muscimol (40 nm) was measured with solubilized receptors precipitated by either a single antibody or a combination of antibodies, as indicated. The calculated values for pairs of antibodies represent the sums of receptors immunoprecipitated by each antibody alone. In this representative experiment, the following values were determined: α5, 31.4%; γ2, 59.4%; γ3, 7.6%; α5γ2, 66.7%; α5γ3, 38.1% of [3H]muscimol binding sites.
Discussion
Pharmacological evidence for α5β3γ2 as a native GABAA receptor isoform.
In a previous study, it was shown that L-655,708 has at least a 50-fold selectivity for α5 versus α1, α2, α3, and α6 subunit-containing receptors and that the tritiated compound [3H]L-655,708 binds rapidly and reversibly to brain membranes, establishing this drug as a specific tool to investigate native α5-containing receptors in more detail (Quirk et al., 1996). Here, the pharmacological characteristics of α5 subunit-containing receptors from rat and human hippocampus have been analyzed with the selective tritiated ligandL-655,708 and compared with those of recombinant human α5β3γ2 or α5β3γ3 GABAA receptors expressed in cell lines.
Scatchard analysis of [3H]L-655,708 binding showed that α5 receptors have similar levels of expression in human and rat hippocampus, as indicated by comparableB max values. The ratB max value (251 fmol/mg of protein) is similar to that measured in a previous study (Quirk et al., 1996) and accounts for 15% of [3H]Ro15–1788 binding sites, in agreement with results from immunoprecipitation experiments (McKernan et al., 1991; Mertens et al., 1993). In human hippocampus, α5 receptors seem to be more abundant, representing 28% of [3H]Ro15–1788 binding sites. Although this value was not determined from saturation analysis, the observed difference is not accounted for by reduced Ro15–1788 affinity for human α5 receptors (Table 2). However, we noticed an important variation in the [3H]L-655,708B max values (coefficient of variation = 0.54), and a larger number of human hippocampal specimens should be investigated to confirm this apparent higher proportion of α5-containing receptors. In addition, the saturation experiment data showed that [3H]L-655,708 has some binding selectivity (9-fold) for α5β3γ2 receptors and shows similar Kd values for this isoform and native receptors in rat and human hippocampus.
The similarity of the pharmacological characteristics of α5β3γ2 and hippocampal α5-containing receptors was further established using several selective compounds for α5β3γ2 receptors (CGS8216, diazepam, L-655,708, and flunitrazepam) or α5β3γ3 receptors (CL218872) (Luddens et al., 1994; Hadinghamet al., 1995). The rank order of the tested compounds matches and extends the reported data for native (McKernan et al., 1991; Quirk et al., 1996) and recombinant (Pritchett and Seeburg, 1990; Luddens et al., 1994) α5-containing receptors and indicates that α5β3γ2 is the major isoform of α5-containing receptors expressed in rat and human hippocampus. These data are also in agreement with results from electrophysiological recordings of cells expressing α5β3γ2L receptors and hippocampal CA1 pyramidal neurons, indicating that this isoform is a native GABAA receptor (Burgard et al., 1996).
Structure of rat hippocampal α5 subunit-containing receptors.
The quantitative immunoprecipitation results clearly indicated that our specific anti-γ3 antibody (Quirk et al., 1994a) did not precipitate any [3H]L-655,708 binding sites from rat hippocampus but precipitated >5% of [3H]Ro15–1788 binding sites. This value is similar (p > 0.31, t test) to the 9.3 ± 1.7% of [3H]muscimol binding sites immunoprecipitated from rat hippocampus and fits with the low level of γ3 subunit expression in rat hippocampus (Herb et al., 1992; Wisden et al., 1992). Furthermore, additive immunoprecipitation experiments similar to those used to demonstrate γ2γ3 coassembly (Quirk et al., 1994a) failed to support an association of γ3 with the α5 subunit. In contrast, measured percentages for the α5γ2 combination correspond almost exactly to theoretical values, indicating that probably all α5 subunits coexist with γ2 subunits to form hippocampal GABAAreceptors. Indeed, α5γ2-containing receptors account for 21.8 ± 2.3% of hippocampal [3H]muscimol binding sites, a value similar to the 19 ± 4% of anti-γ2-immunoprecipitated [3H]L-655,708 binding sites and the [3H]L-655,708/[3H]Ro15–1788 ratio (15 ± 2%) (p > 0.40, one-way analysis of variance). Although the widely accepted stoichiometry of GABAA receptors is 2α2β1γ (for review, seeMcKernan and Whiting, 1996), quantitative immunoprecipitation and Western blot analysis have revealed the coexistence of γ2 and γ3 in approximately 7% of rat brain GABAA receptors (Quirk et al., 1994a). The pharmacological characteristics of these γ2γ3-containing isoforms have not been analyzed, and the possibility that the γ2 subunit is pharmacologically predominant over the γ3 subunit, thus masking the detection of α5β3γ2γ3 complexes, cannot be excluded. Such a predominant effect has been shown for the α1 subunit over the α3 subunit in native cortical GABAA receptors (Araujo et al., 1996), but not over the α6 subunit in cerebellar receptors (Khan et al., 1996).
The presence of low levels of [3H]L-655,708 binding associated with anti-α1-, anti-α2-, and anti-α3-immunoprecipitated receptors suggested the existence of receptors with mixed contents of α subunits. By combining the estimated amounts of α1 (43%), α2 (18%), and α3 (17%) in rat hippocampus (McKernan and Whiting, 1996) and the [3H]L-655,708 data, it is possible to estimate that α5αX=1,2,3receptors account for ∼15% of total α5 subunit-containing receptors in rat hippocampus. Although these isoforms are of low abundance, it seems that the pharmacological characteristics of the α5 subunit predominate, as judged by the binding of the α5-selective ligand [3H]L-655,708. A contribution of each α subunit to the overall receptor pharmacology has been shown in α1α6- and α1α3-containing receptors, with more or less predominance of one subunit over the other (Khan et al., 1996; Araujo et al., 1996).
In a recent study, Fritschy et al. (1997) showed that levels of α5, β2/3, and γ2 subunit expression remain unaffected in γ3-deficient mutant mice, suggesting the coassembly of these proteins to form native GABAAreceptors. The pharmacological and biochemical data reported here support and strengthen such a conclusion, because they demonstrate a preferential association of the α5 subunit with the γ2 subunit in both rat and human hippocampus and establish α5β2/3γ2 as a native hippocampal GABAA receptor isoform.
Acknowledgments
We gratefully acknowledge the support of the Molecular Biology Department.
Footnotes
- Received May 14, 1998.
- Accepted June 26, 1998.
-
Send reprint requests to: Dr. Cyrille Sur, Department of Biochemistry, Merck Sharp and Dohme Research Laboratories, Terlings Park, Eastwick Road, Harlow, Essex, CM20 2QR, UK. E-mail:crrille_sur{at}merck.com
Abbreviation
- GABA
- γ-aminobutyric acid
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}