


Abstract
Pituitary adenylate cyclase-activating polypeptide (PACAP) is a 38-amino acid peptide that was first isolated from ovine hypothalamic extracts on the basis of its ability to stimulate cAMP formation in anterior pituitary cells. PACAP belongs to the vasoactive intestinal polypeptide (VIP)-glucagon-growth hormone releasing factor-secretin superfamily. The sequence of PACAP has been remarkably well conserved during the evolution from protochordate to mammals, suggesting that PACAP is involved in the regulation of important biological functions. PACAP is widely distributed in the brain and peripheral organs, notably in the endocrine pancreas, gonads, and respiratory and urogenital tracts. Characterization of the PACAP precursor has revealed the existence of a PACAP-related peptide whose activity remains unknown. Two types of PACAP binding sites have been characterized. Type I binding sites exhibit a high affinity for PACAP and a much lower affinity for VIP whereas type II binding sites have similar affinity for PACAP and VIP. Molecular cloning of PACAP receptors has shown the existence of three distinct receptor subtypes, the PACAP-specific PAC1 receptor, which is coupled to several transduction systems, and the two PACAP/VIP-indifferent VPAC1 and VPAC2 receptors, which are primarily coupled to adenylyl cyclase. PAC1 receptors are particularly abundant in the brain and pituitary and adrenal glands whereas VPAC receptors are expressed mainly in the lung, liver, and testis. The wide distribution of PACAP and PACAP receptors has led to an explosion of studies aimed at determining the pharmacological effects and biological functions of the peptide. This report reviews the current knowledge concerning the multiple actions of PACAP in the central nervous system and in various peripheral organs including the endocrine glands, the airways, and the cardiovascular and immune systems, as well as the different effects of PACAP on a number of tumor cell types.
I. Introduction
The secretory activity of the adenohypophysis is regulated by aminergic (mainly dopaminergic) and peptidergic hypothalamic neurons (Elde and Hökfelt, 1979; Stumpf and Jennes, 1984; Ju et al., 1991). Five neuropeptides have been isolated from ovine and porcine hypothalamic extracts, or from a human pancreatic tumor, and characterized by the groups of Roger Guillemin, Andrew Schally, and Willy Vale, based on their ability to either stimulate or inhibit the secretion of anterior pituitary hormones: thyrotropin-releasing hormone (TRH; Boler et al., 1969; Burgus et al., 1969), gonadotropin-releasing hormone (GnRH; Amoss et al., 1971; Matsuo et al., 1971), somatostatin (Brazeau et al., 1973; Esch et al., 1980;Böhlen et al., 1981), corticotropin-releasing factor (CRF; Vale et al., 1981), and growth hormone-releasing factor (GRF; Guillemin et al., 1982; Rivier et al., 1982b). All of these hypophysiotropic neurohormones are synthesized in hypothalamic neurons, whose axons project toward the median eminence, and are transported to the anterior pituitary by the capillaries of the portal system. Another common feature of these hypothalamic neurohormones is that they are generally widely distributed in the central nervous system (CNS) and in peripheral organs, and that they exert a large array of biological activities in addition to their hypophysiotropic actions. After the primary structure of GRF had been determined in 1982, it was commonly thought that all major hypophysiotropic neurohormones had been identified. However, the subsequent characterization of other neuropeptides capable of regulating the activity of anterior pituitary cells, such as pituitary adenylate cyclase-activating polypeptide (PACAP; Miyata et al., 1989) and prolactin (PRL)-releasing peptide (Hinuma et al., 1998), has shown that this view was incorrect.
PACAP has been originally isolated from an extract of ovine hypothalamus on the basis of its ability to stimulate cAMP formation in rat pituitary cells (Miyata et al., 1989). Hypothalamic neurons containing PACAP project toward the median eminence and terminate in the vicinity of the capillary loops of the hypothalamo-pituitary portal system. Like other hypophysiotropic neurohormones, PACAP is contained in extra-hypothalamic neurons as well as in numerous peripheral tissues. Consistent with its widespread distribution, PACAP has been found to exert pleiotropic effects including modulation of neurotransmitter release, vasodilation, bronchodilation, activation of intestinal motility, increase of insulin and histamine secretion, as well as stimulation of cell multiplication and/or differentiation.
II. PACAP
A. Discovery of PACAP
To isolate novel hypophysiotropic neuropeptides, the group of Arimura has screened fractions from an extract of 4300 ovine hypothalamus by monitoring their stimulatory effect on adenylyl cyclase activity in cultured rat anterior pituitary cells. Using this approach, they have isolated in pure form a peptide, found to markedly increase cAMP formation, that they termed pituitary adenylate cyclase-activating polypeptide. Characterization of the peptide revealed that it comprises 38 amino acid residues and is C-terminally α-amidated (Miyata et al., 1989). Two years later, the primary structure of this 38-amino acid form of PACAP (PACAP38) was determined in the European green frog Rana ridibunda, a species that diverged from the line leading to mammals some 280 million years ago (Chartrel et al., 1991; Hoyle, 1998). Frog PACAP38 appears to contain only one amino acid substitution (Val35 → Ile), which may be accomplished by the exchange of a single nucleotide in the cDNA sequence (Chartrel et al., 1991). The sequence of PACAP38 comprises an internal cleavage-amidation site (Gly28-Lys29-Arg30), suggesting that the PACAP precursor can generate a 27-residue α-amidated polypeptide (PACAP27). Consistent with this hypothesis,Miyata et al. (1990) have isolated from the ovine hypothalamus another fraction capable of stimulating adenylyl cyclase activity in adenohypophysial cells that, on characterization, happened to correspond to the N-terminal 27-amino acid sequence of PACAP38. Thus it appears that the structure of the biologically active region of PACAP, corresponding to the PACAP27 sequence, has been totally preserved during evolution, from amphibians to mammals. The sequence of PACAP27 shows 68% identity with vasoactive intestinal polypeptide (VIP), identifying PACAP as a member of the VIP-glucagon-GRF-secretin superfamily of structurally related peptides (Fig.1; Campbell and Scanes, 1992; Segre and Goldring, 1993).

Amino acid sequences of the different members of the PACAP-VIP-GRF-glucagon superfamily in human. –, amino acids identical with those of PACAP38. Adapted from Kieffer and Habener, 1999.
B. Secondary Structure of PACAP
Conformational analysis of PACAP27 by two-dimensional NMR and circular dichroism spectroscopy has shown an initial disordered N-terminus sequence of eight amino acid residues followed by a region, from amino acid residues 9 to 24, that consists of four distinct domains (Inooka et al., 1992). The first domain, encompassing residues 9 to 12, forms a β-turn-like conformation whereas the three others are composed of distinct helical regions that extend from residues 12 to 14, 15 to 20, and 22 to 24, respectively. The conformation of PACAP38 mirrors that of PACAP27 in its N-terminal region whereas the C-terminal segment exhibits a short helix attached by a flexible hinge to the 1–27 region (Wray et al., 1993). The biological importance of the three structural domains of PACAP38 has been investigated using truncated PACAP analogs (see Section III, D).
The three-dimensional structure of PACAP exhibits substantial similarities with those of other members of the VIP/glucagon family (Braun et al., 1983; Gronenborn et al., 1987; Wray et al., 1993). In particular, both PACAP27 and VIP possess two helices separated by a disordered region, but the position of the first α-helix of PACAP27 is shifted by two residues toward the C-terminus, and the conformation of the second helix of PACAP27 is closer to an α-helix than that of VIP. These minor conformational differences between PACAP27 and VIP may contribute to the selectivity of the peptides for their receptors (Inooka et al., 1992).
C. Structure of the PACAP Precursor and Post-Translational Processing
The cDNA encoding the PACAP precursor has been characterized in several vertebrate species (Ogi et al., 1990; Ohkubo et al., 1992;Arimura and Shioda, 1995; Okazaki et al., 1995) and in a protochordate, the ascidian Chelyosoma productum (McRory and Sherwood, 1997). In humans, the cDNA encodes a 176-amino acid prepro-protein, which comprises a 24-amino acid signal peptide (Hosoya et al., 1992). In all mammalian species studied so far, the sequence of PACAP38 is located in the C-terminal domain of the precursor (Fig.2). The cDNA sequences of humans (Ohkubo et al., 1992), sheep (Kimura et al., 1990), rat (Ogi et al., 1990), and mouse prepro-PACAP (Okazaki et al., 1995) have revealed the existence of a 29-amino acid peptide delimited by basic residues at its N- and C-terminal extremities, located upstream of PACAP38 (Fig. 2). This peptide, which exhibits moderate structural homology with PACAP27, has been termed PACAP-related peptide (PRP) (Ogi et al., 1990; Wray et al., 1995; Hoyle, 1998). In mammals, the overall organization of the PACAP precursor exhibits strong similarities with that of the VIP precursor (Fig. 2). In particular, the VIP precursor encompasses a VIP-related peptide, called peptide histidine-methionine (PHM) amide in humans (Itoh et al., 1983; Bodner et al., 1985; Christophe et al., 1989) or peptide histidine-isoleucine (PHI) amide in sheep (Bounjoua et al., 1991), rat (Nishizawa et al., 1985), mouse (Lamperti et al., 1991), and chicken (McFarlin et al., 1995), which possesses moderate amino acid identity with VIP. The degree of similarity between PACAP27 and PRP (22%) or VIP and PHI (37%) is less than that between PACAP and VIP (68%) or PRP and PHI (44%), respectively. Thus it is assumed that intragenomic duplication of a VIP/PACAP ancestor sequence has occurred before duplication of the whole ancestor gene (Ohkubo et al., 1992). A proposed model describing the evolutionary process leading to the generation of distinct precursors for PACAP, VIP, glucagon, GRF, and secretin in mammals is presented in Fig.3. In submammalian vertebrates and the tunicate Chelyosoma productum, the PACAP precursor comprises both GRF and PACAP (Fig. 3) (Parker et al., 1993; McRory et al., 1995,1997; McRory and Sherwood, 1997; Alexandre et al., 2000) (See section II, I).

Schematic representation of the human PRP/PACAP and PHM/VIP precursors. The general organization of the two precursors is presented and the sequences of PRP and PHM as well as PACAP and VIP have been aligned. The conserved amino acids are indicated in black and the percentage of amino acid identity between PRP and PHM as well as PACAP27 and VIP are indicated. SP, signal peptide.

Hypothetical schemes depicting the evolutionary history of the PACAP/VIP/glucagon/GRF/secretin gene family. Adapted from Ohkubo et al., 1992.
In mammals, the primary structure of the PACAP precursor reveals the existence of seven mono- or dibasic residues that can be cleaved by various prohormone convertases (PCs) including PC1, PC2, PC4, PC5, PC7, furine, and PACE4 (Seidah et al., 1994, 1998). In the rat, cleavage at three dibasic sites, i.e., Arg79-Arg80, Lys129-Arg130, and Arg170-Arg171, generates a large intermediate precursor of PRP (big PRP) and a glycine-extended form of PACAP38 (Fig. 4). Cleavage at the single Arg110, followed by hydrolysis of this C-terminal Arg residue by carboxypeptidases E, H, or M, generates PRP (Rouillé et al., 1995). The Gly169 residue is used by peptidyl glycine α-amidating monooxygenase (Eipper et al., 1992a,b) for the amidation of the Lys168 residue at the C-terminal extremity of PACAP38. Finally, the tripeptide Gly158-Lys159-Arg160can be cleaved to generate the α-amidated PACAP27 isoform (Fig. 4). Processing of the PACAP precursor has been studied in Chinese hamster ovary (CHO)-K1 cells transfected with the human PACAP cDNA (Okazaki et al., 1992). Characterization of the various peptides secreted in the incubation medium by HPLC combined with radioimmunoassay (RIA) detection has confirmed that processing of the PACAP precursor actually yields to the formation of PACAP38, PACAP27, and PRP (Okazaki et al., 1992).

Schematic representation of the post-translational processing of the rat PACAP precursor. The nature and allocation of each cleavage and amidation site is specified. PAM, peptidyl glycine α-amidating monooxygenase. SP, signal peptide.
In the rat hypothalamus, PC1 and/or PC2 are intensively expressed in nuclei enriched with PACAP-immunoreactive neurons, supporting the hypothesis that these two endopeptidases could be involved in the processing of the PACAP precursor (Köves et al., 1994a; Zheng et al., 1994; Dong et al., 1997). Cotransfection experiments in GH4C1 cells have confirmed that both PC1 and PC2 can actually process the rat PACAP precursor to generate mature PACAP38 and PACAP27 (Li et al., 1999). In the testis, where PACAP is particularly abundant, PC4 can process the PACAP precursor to generate both PACAP38 and PACAP27 (Li et al., 1998).
D. The PACAP Gene
The gene encoding PACAP has been cloned in humans (Hosoya et al., 1992) and mouse (Yamamoto et al., 1998). The overall architecture of the two genes is similar, with the exception of the 5′-untranslated region of the mouse gene, which encompasses two exons as a result of alternative splicing of the transcription initiation domain. The humanPACAP gene is composed of five exons, the sequence of PRP being encoded by exon 4 and that of PACAP by exon 5 (Fig.5). Northern blot analysis has revealed the presence of a 3-kb PACAP mRNA in the rat hypothalamus (Hosoya et al., 1993; Hannibal et al., 1995a). A shorter transcript with a truncated 5′-untranslated region has been characterized in the rat testis (Hurley et al., 1995). Similarly, shorter PACAP mRNA has been found in the mouse, bovine, and human testis (Hurley et al., 1995). It has also been reported that another short PACAP transcript is produced in sympathetic neurons (Harakall et al., 1998).

Organization of the human PACAP gene and PACAP mRNA. The five exons are boxed and numbered. The untranslated regions of exons 1 and 5 are denoted by dashed lines. Exon domains encoding PRP and PACAP are hatched. Arrows indicate the locations of binding sites for potential transcriptional factors. CRE, cAMP response element; Inr-like, initiator-like element; P1, promotor region 1; P2, promotor region 2; TRE, 12-O-tetradecanoylphorbol 13-acetate response element. GHF-1, growth hormone factor 1.
The promoter sequence of the PACAP gene (about 400 bp) comprises two regions, termed P1 and P2 (Fig. 5), which correspond, respectively, to an initiator-like sequence and a CT-rich domain with GC boxes (Jankowski and Dixon, 1987; Ohkubo et al., 1994). Surprisingly, the promoter region of the human PACAP gene does not contain any apparent TATA or CAAT box, which are normally required for accurate initiation of transcription (Hampsey, 1998). In contrast, the PACAP promoter possesses two cAMP-response-like elements, a 12-O-tetradecanoylphorbol 13-acetate response element and a pair of sequences homologous to the consensus sequence for pituitary-specific factor growth hormone factor 1-binding sites, which are known to play a role in the tissue-specific expression of growth hormone (GH) (Bodner et al., 1988; Dolle et al., 1990; Castrillo et al., 1991). Investigation of the promoter activity has revealed that PACAP is constitutively expressed and that transcription of thePACAP gene can be enhanced by cAMP, 12-O-tetradecanoylphorbol 13-acetate, and even by PACAP itself (Suzuki et al., 1994a; Hashimoto et al., 2000).
The structural organization of the PACAP gene is similar to that of the VIP gene (Lamperti et al., 1991) andGRF gene (Mayo et al., 1985), confirming that all three genes originate from a common ancestral sequence through gene duplication (Fig. 3). In humans, the PACAP gene has been localized by Southern blotting and in situ hybridization to the P11 region of chromosome 18. This region is associated with holoprosencephaly, the most common hereditary developmental defect of the forebrain in humans, suggesting that PACAP might be involved in the control of brain development (Hosoya et al., 1992; Chang et al., 1993;Golden, 1998).
E. Distribution of PACAP in the CNS
Soon after the characterization of PACAP, the distribution of the peptide was determined in the brain of mammals (Arimura et al., 1991;Köves et al., 1991; Vigh et al., 1991; Kivipelto et al., 1992;Ghatei et al., 1993) and amphibians (Yon et al., 1992). The distribution of PACAP-immunoreactive cells and fibers in the rat brain was schematically presented in a previous review (Gonzalez et al., 1998). In rat, RIA measurements have revealed that the highest concentrations of PACAP occur in the hypothalamic area (Arimura et al., 1991; Ghatei et al., 1993). Reversed-phase HPLC analysis showed that PACAP38 is by far the predominant form, PACAP27 representing less than 10% of the total peptide content in brain tissue (Arimura et al., 1991; Ghatei et al., 1993; Masuo et al., 1993; Hannibal et al., 1995a;Piggins et al., 1996).
The mapping of PACAP-expressing neurons has been investigated by in situ hybridization and immunocytochemistry (Table1). In the rat hypothalamus, PACAP-immunoreactive neurons are primarily located in the parvo- and magnocellular neurons of paraventricular and supraoptic nuclei (Köves et al., 1991, 1994b; Kivipelto et al., 1992; Ando et al., 1994; Kimura et al., 1994; Hannibal et al., 1995a,b; Piggins et al., 1996). PACAP mRNA is expressed in the paraventricular and arcuate nuclei (Hannibal et al., 1995b; Murase et al., 1995). A dense accumulation of PACAP-immunoreactive fibers is found in the internal zone of the median eminence and in the vicinity of the capillaries of the hypothalamo-hypophysial portal system (Köves et al., 1990,1991; Kivipelto et al., 1992; Tamada et al., 1994; Hannibal et al., 1995a,b; Mikkelsen et al., 1995). Quantification of PACAP by RIA has shown that the concentration of the peptide in the rat portal blood is significantly higher than in the peripheral blood, indicating that PACAP released by hypothalamic nerve terminals is actually transported to the pituitary (Dow et al., 1994). Regional distribution studies revealed that significant amounts of PACAP38 are also found in extrahypothalamic regions, including the substantia nigra, nucleus accumbens, septum, globus pallidus, cerebral piriform cortex, and pons (Ghatei et al., 1993; Masuo et al., 1993). In the limbic system, PACAP-like immunoreactive fibers are detected in the amygdaloid complex and in the mediodorsal and paraventricular nuclei of the thalamus (Köves et al., 1991; Masuo et al., 1993; Takahashi et al., 1994;Palkovits et al., 1995). In the lateral septum area, a dense network of immunoreactive fibers innervates blood vessels (Köves et al., 1991). In situ hybridization has revealed the presence of scattered PACAP-expressing cell bodies in the cingulate and frontal cortex (Mikkelsen et al., 1994). PACAP and its mRNA also have been detected in the cerebellum (Ghatei et al., 1993; Mikkelsen et al., 1994; Takahashi et al., 1994; Hannibal et al., 1995a; Nielsen et al., 1998a). Specifically, PACAP-like immunoreactivity (PACAP-LI) is localized in the soma and dendrites of Purkinje cells, whose axons directly contact granule cells (Nielsen et al., 1998a). In the medulla oblongata, the majority of perikarya exhibiting PACAP-LI are found in the commissural and medial subnuclei of the solitary nucleus, the dorsal motor vagal nucleus, the nucleus ambiguous, the ventrolateral medulla, the ventral medullary surface, and the caudal raphe nuclei, supporting the hypothesis that PACAP may act as a regulator of visceral functions (Legradi et al., 1994). In the spinal cord, PACAP mRNA is expressed in a subpopulation of sensory neurons of the dorsal root ganglia (Mulder et al., 1994), and numerous PACAP-immunoreactive fibers are found in the superficial layer of the dorsal horns (Moller et al., 1993; Dun et al., 1996a).
Localization and relative abundance of PACAP mRNA and PACAP-like immunoreactivity in the rat brain
The location of PACAP-containing neurons also has been investigated in the CNS of nonmammalian vertebrates, including birds (Peeters et al., 1998), amphibians (Yon et al., 1992, 1993b), and fishes (Matsuda et al., 1997a,b; Montéro et al., 1998). Globally, the distribution of PACAP-immunoreactive cells exhibits a high degree of similarity with that of mammals. In particular, in the brain of the frog Rana ridibunda, prominent groups of PACAP-containing neurons are located in the hypothalamus, i.e., in the anterior preoptic area, the ventral magnocellular preoptic nucleus, the suprachiasmatic nucleus, the ventral hypothalamic nucleus, and the posterior tubercle (Yon et al., 1992). Similarly, in the primitive teleost fish Anguilla anguilla, PACAP-containing neurons are primarily located in the parvo- and magnocellular subdivisions of the preoptic nucleus (Montéro et al., 1998).
The distributions of PACAP and VIP in the CNS are substantially different (Masuo et al., 1993). For instance, in the thalamus a few VIP fibers were found running up the wall of the third ventricle whereas a dense network of PACAP fibers was observed in the central thalamic nuclei (Köves et al., 1991). In the bed nucleus of stria terminalis, PACAP fibers appear to surround unstained, round-shaped neuronal cell bodies, whereas the VIP fibers are homogeneously distributed. PACAP-immunoreactive fibers are also found in the lateral septum of the hypothalamus where only a few VIP fibers are observed (Köves et al., 1991). In the magnocellular neurons, PACAP but not VIP is colocalized with oxytocin (Köves et al., 1994b). In the brainstem, VIP-LI is present in the mesencephalic periaqueductal gray and the dorsal and linear raphe nuclei whereas PACAP-LI is abundant in the paraventricular nucleus (PVN) and the dorsal vagal complex. The bed nucleus of the stria terminalis contains a very high concentration of PACAP and VIP-LI but no double-labeled cells have been detected (Kozicz et al., 1997). In contrast, both PACAP and VIP-immunoreactive fibers appear to innervate the wall of cerebral blood vessels (Jansen-Olesen et al., 1994).
F. Distribution of PACAP in Peripheral Organs
In peripheral tissues, as in the brain, PACAP38 is by far the major molecular form but the proportions of PACAP27 and PACAP38 vary between the different organs (Arimura et al., 1991). For instance, in the colon, PACAP27 represents 30% of the total immunoreactivity whereas, in the testis, PACAP27 is hardly detectable (Arimura et al., 1991). The occurrence of different proportions of the two peptides in various tissues can be likely ascribed to the existence of different sets of PC enzymes.
The presence of PACAP mRNA and PACAP has been detected in most endocrine glands (Table 2). In particular, PACAP is found in the different lobes of the pituitary gland (Arimura and Shioda, 1995; Rawlings and Hezareh, 1996;Arimura, 1998). In the anterior pituitary, PACAP is observed in a subpopulation of gonadotrope cells (Mikkelsen et al., 1995; Köves et al., 1998). In the ventral part of the neural lobe, PACAP is contained in nerve fibers with large terminal boutons (Mikkelsen et al., 1995). At the ultrastructural level, PACAP-LI appears to be located in dense core granules contained in neurosecretory fibers (Kimura et al., 1994). PACAP-immunoreactive elements are also found in the gonads (Shioda et al., 1994), adrenal (Arimura et al., 1991), parathyroid (Luts and Sundler, 1994), and endocrine pancreas (Table 2;Arimura and Shioda, 1995; Love and Szebeni, 1999). In rat, the highest amounts of PACAP are found in the testis. In fact, the concentration of PACAP in the testis is higher than in the whole brain and exceeds the concentration of any other known peptides (Arimura et al., 1991). In situ hybridization studies have shown that PACAP mRNA is present in germ cells and not in Sertoli or Leydig cells (Shioda et al., 1994). Electron microscopic studies have revealed that PACAP is located in acrosoma caps and granules of primary spermatocytes but the peptide has not been observed in mature spermatids (McArdle, 1994;Shioda et al., 1994; Hannibal and Fahrenkrug, 1995). In the ovary, the concentration of PACAP is much lower than in the testis, and the peptide appears to be contained in nerve fibers (Steenstrup et al., 1995). Intense expression of PACAP mRNA has also been observed in the granulosa cells of preovulatory follicles (Ko et al., 1999). The adrenal gland contains a high concentration of PACAP (Arimura et al., 1991; Watanabe et al., 1992; Ghatei et al., 1993). In mammals, PACAP is found in the adrenal medulla (Shiotani et al., 1995), where it is contained both in chromaffin cells (Holgert et al., 1996) and in fibers (Frödin et al., 1995; Moller and Sundler, 1996). In contrast, in the frog adrenal gland, PACAP-LI is restricted to nerve fibers that contact either chromaffin cells or steroid-producing cells (Yon et al., 1993a). Similarly, in mammals, the parathyroid gland and the intrapancreatic ganglia are innervated by PACAP-containing fibers (Luts and Sundler, 1994; Filipsson et al., 1998a; Love and Szebeni, 1999).
Localization and relative abundance of PACAP mRNA and PACAP-like immunoreactivity in rat peripheral tissues
Large amounts of PACAP-LI are found in all parts of the gastrointestinal tract (Arimura et al., 1991; Hauser-Kronberger et al., 1992; Ghatei et al., 1993; Mao et al., 1998; Vincze et al., 1999). The presence of PACAP-immunoreactive cell bodies has been observed in the myenteric ganglia throughout the gastrointestinal tract, and the existence of intrinsic neurons has been confirmed by in situ hybridization (Shen et al., 1992; Hannibal et al., 1998). Numerous PACAP-containing nerve fibers have been visualized along the circular muscle fibers and in the longitudinal smooth muscle layer of the esophagus (Uddman et al., 1991a; Köves et al., 1993; Olsson and Holmgren, 1994). PACAP-LI has also been detected in various exocrine glands of the alimentary canal, e.g., the parotid and submandibular glands, the liver, and the exocrine pancreas (Arimura et al., 1991;Fridolf et al., 1992; Moller et al., 1993; Luts and Sundler, 1994). In the urinary bladder, networks of PACAP-immunoreactive fibers are found in the vicinity of blood vessels (Moller et al., 1993; Fahrenkrug and Hannibal, 1998). In the airways, PACAP-immunoreactive fibers innervate smooth muscle bundles and blood vessels in the trachea as well as small bronchioles in the lung (Cardell et al., 1991; Uddman et al., 1991b;Hauser-Kronberger et al., 1996; Shigyo et al., 1998). In the immune system, PACAP is expressed in various lymphoid tissues including the thymus, spleen, and duodenal mucosa (Gaytan et al., 1994), and in peritoneal macrophages (Pozo et al., 1997). The occurrence of PACAP mRNA has been demonstrated in the superior cervical ganglion (Nogi et al., 1997b). Depolarization of these neurons stimulates the release of PACAP27 and PACAP38 and causes a concomitant increase of PACAP mRNA and peptide (Brandenburg et al., 1997). A few PACAP-positive perikarya are also present in the sphenopalatine and otic ganglia (Uddman et al., 1991b, 1999). In the eye, PACAP-LI is present in fibers innervating the iris sphincter and in cell bodies scattered in the ciliary ganglia (Wang et al., 1995; Elsas et al., 1997; Olianas et al., 1997; Samuelsson-Almen and Nilsson, 1999) and in fibers of the ganglion cell layer of the retina (Hannibal et al., 1997; Seki et al., 1997).
In peripheral organs, in contrast to the CNS, PACAP and VIP often appear to be coexpressed by the same cells. For instance, colocalization of PACAP and VIP has been demonstrated in nerve fibers and cell bodies in the human and sheep esophageal sphincter (Uddman et al., 1991a; Ny et al., 1995), in the human and chicken gut (Sundler et al., 1992), and in the ovine respiratory tract (Uddman et al., 1991b). Nerve fibers containing both PACAP and VIP are also found in other tissues, notably in the parathyroid glands of cat and sheep (Luts and Sundler, 1994), and in the gill arch of the goldfish Carassius auratus (De Girolamo et al., 1998).
G. PACAP in Tumor Cells
The PACAP gene is differentially expressed in brain tumors. PACAP mRNA is present in most gliomas but is detected in only one-fifth of meningiomas (Vertongen et al., 1995a). PACAP mRNA and PACAP-LI are abundant in human neuroblastomas (Suzuki et al., 1993; Takahashi et al., 1993a; Vertongen et al., 1997a; Waschek et al., 1997). Double-staining experiments have demonstrated that PACAP and VIP are colocalized and intensely expressed in most pancreatic carcinoma, neuroblastoma, and pheochromocytoma tumors (Fahrenkrug et al., 1995). VIP has been reported to exert an autocrine stimulation of neuroblastoma cell growth and differentiation. The presence of PACAP suggests that it could also control neuroblastoma cell tumor proliferation (O'Dorisio et al., 1992; Pence and Shorter, 1992). Most pituitary tumors contain large amounts of PACAP. Because pituitary cells are programmed to proliferate in response to cAMP (Lin et al., 1992), it is conceivable that in pituitary adenomas, PACAP contributes to tumorigenesis (Spada et al., 1996). Overexpression of PACAP has also been reported in ovarian tumors (Odum and Fahrenkrug, 1998) and in pheochromocytomas (Takahashi et al., 1993b).
H. Ontogenesis of PACAP
The evolution of the content of PACAP during development has been studied in detail in the CNS of rodents (Shuto et al., 1996; Waschek et al., 1998; Skoglösa et al., 1999b,c). In the mouse embryo, PACAP mRNA is present in the brain as early as embryonic day 9.5 (E9.5) (Shuto et al., 1996; Waschek et al., 1998), and the mRNA level increases during the prenatal period to reach a maximum at birth. In situ hybridization histochemistry revealed that the PACAPgene is widely expressed in the neural tube of the mouse at E10.5 (Shuto et al., 1996; Waschek et al., 1998). PACAP mRNA is found in differentiating neurons, suggesting that PACAP may control proliferation or differentiation of neuroblasts during neural tube development. PACAP is readily measurable by RIA in the rat brain at E14 (Masuo et al., 1994; Tatsuno et al., 1994). Immunoreactive nerve fibers are observed in the spinal cord and ganglia at E16 (Nielsen et al., 1998b). In the septum and hypothalamus, the content of PACAP increases gradually from birth to postnatal day 60 (P60). In the cortex, hippocampus, thalamus, and midbrain, PACAP levels increase more rapidly from P10 to P20 and reach a plateau at P30 (Masuo et al., 1994). In the striatum and cerebellum, the content of PACAP is very high at birth and during the first postnatal weeks and then decreases gradually from P20 to adulthood. In the developing rat cerebellum, PACAP is expressed in Purkinje cells (Nielsen et al., 1998a; Skoglösa et al., 1999b), which are known to regulate the survival of granule cells.
The ontogenesis of PACAP has also been described in the brain of the frog Rana ridibunda (M. Mathieu, L.Yon, I. Charifou, M. Trabucchi, M. Vallarino, C. Pinelli, R.K. Rastogi and H.Vaudry, submitted). PACAP-immunoreactive neurons are found soon after hatching (stages IV-VII of development; Taylor and Kollros, 1946) in the dorsal thalamus, and appear later (stages VII-IX) in the dorsal and ventral infundibular nuclei of the hypothalamus. PACAP-immunoreactive fibers are seen in the median eminence during the premetamorphic period (stages XIII-XVIII), suggesting that PACAP could be involved in the activation of the pituitary-thyroid axis, which is required for the onset of metamorphosis (Tata, 1998). Reversed-phase HPLC analysis combined with RIA detection indicates that PACAP38 is, by far, the predominant molecular form present in the frog brain at all developmental stages (M. Mathieu, L.Yon, I. Charifou, M. Trabucchi, M. Vallarino, C. Pinelli, R.K. Rastogi and H.Vaudry, submitted).
I. Phylogenetic Evolution of PACAP
The primary structure of PACAP has been totally conserved among those mammalian species yet studied, i.e., human (Ohkubo et al., 1992), sheep (Miyata et al., 1989), rat (Ogi et al., 1990), and mouse (Okazaki et al., 1995). The sequence of PACAP has now been determined in several representative species of nonmammalian vertebrates, including the chicken Galus domesticus (McRory et al., 1997), the frogRana ridibunda (Chartrel et al., 1991), the salmonOncorhynchus nerka (Parker et al., 1993), the catfishClarias macrocephalus (McRory et al., 1995), and the tunicate Chelyosoma productum (McRory and Sherwood, 1997) (Fig. 6). A partial sequence of PACAP that is identical with the first 28 amino acids of mammalian PACAP38 has also been characterized in the lizard Gila monstersalivary gland (Pohl and Wank, 1998), and the presence of PACAP-LI has been documented in the brain and ovary of the crested newt,Triturus carnifex (Gobbetti et al., 1997). The primary structure of the 1–27 region of PACAP, which is responsible for the biological activity of the peptide, has been fully conserved in lizard, frog, salmon, and catfish, whereas the PACAP27 sequences of the chicken and stargazer exhibit only one amino acid substitution (Fig.6). In contrast, the C-terminal portion of PACAP, which is not required for the biological activity of the peptide, is more variable (Fig. 6). Globally, the sequence of PACAP has been better preserved than that of VIP (Chartrel et al., 1995) and far more conserved than that of GRF across vertebrates (M. Montéro, L.Yon, D. Kikuyama, S. Dufour and H.Vaudry, submitted). The fact that evolutionary pressure has acted to strongly preserve the bioactive sequence of PACAP indicates that the peptide must exert important physiological functions. In support of this notion, a PACAP-like peptide has been identified in the insectDrosophila melanogaster (Feany and Quinn, 1995), and this peptide has been found to modulate ionic conductances at the neuromuscular junction (Zhong, 1995; Zhong and Pena, 1995).

Comparison of the amino acid sequences of PACAP from various vertebrate species and a protochordate. Percentages indicate amino acid identity between PACAP38 from different nonmammalian vertebrates and mammalian PACAP38, and between lizard or tunicate PACAP27 and mammalian PACAP27. –, amino acids identical with those of human PACAP. The potential cleavage-amidation sites are underlined.
Two different genes for PACAP are present in the tunicateChelyosoma productum (Fig. 6; McRory and Sherwood, 1997). Each of these genes encodes both PACAP and a GRF-like peptide (Fig. 3). Nucleotide sequence similarities suggest that the two tunicatePACAP genes arose from exon duplication followed by gene duplication. In salmon, a cDNA that encodes both PACAP and a GRF-like peptide has been characterized (Parker et al., 1993). A cDNA encoding both PACAP and GRF-like peptide has also been cloned in the catfishClarias macrocephalus (McRory et al., 1995), frog Rana ridibunda (Alexandre et al., 2000), and chicken Gallus domesticus (McRory et al., 1997). In salmon, catfish, and chicken, alternative splicing of the primary transcript generates a shorter precursor that contains only PACAP (Parker et al., 1993; McRory et al., 1995, 1997). In contrast to all submammalian species investigated so far, in mammals, GRF and PACAP precursors are encoded by two distinct genes (Mayo et al., 1985; Hosoya et al., 1992). Based on primary sequence homologies among existing peptides of the GRF superfamily (Fig. 1), it is possible to construct a hypothetical evolution tree of these genes (Campbell and Scanes, 1992). The organization of the mammalian prepro-GRF and prepro-PACAP cDNAs suggests that the two genes arose from duplication of an ancestral gene with subsequent exon loss (Fig. 3; Parker et al., 1997). Within thePACAP-VIP-glucagon-GRF-secretin gene superfamily, thePACAP gene appears to be closely related to theVIP one (Ogi et al., 1990). Furthermore, by comparison of the peptide sequences and geological record, one can predict the changes that have occurred during the evolution of the VIP-glucagon-GRF-secretin superfamily. According to these chronological analyses, duplication of a common ancestral gene yielding to thePRP/PACAP and PHI/VIP genes may have occurred some 750 million years ago (Campbell and Scanes, 1992).
III. The PACAP Receptors
A. Pharmacological Characterization of PACAP Receptors
Two classes of PACAP binding sites have been characterized on the basis of their relative affinities for PACAP and VIP (Table3). Type I binding sites, which have been originally characterized in the anterior pituitary and hypothalamus using 125I-PACAP27 as a radioligand, exhibit high affinity for PACAP38 and PACAP27 (K d ≈ 0.5 nM) and much lower affinity for VIP (K d > 500 nM) (Cauvin et al., 1990; Gottschall et al., 1990, 1991; Lam et al., 1990;Suda et al., 1992). Type II binding sites, which are abundant in various peripheral organs including the lung, duodenum, and thymus, possess similar affinity for PACAP and VIP (K d ≈ 1 nM) (Gottschall et al., 1990; Lam et al., 1990). Subtle differences in the ability of PACAP38 and PACAP27 to displace 125I-PACAP27 from its recognition sites in the CNS suggest the existence of two subsets of type I binding sites (Cauvin et al., 1991; Robberecht et al., 1991b). Similarly, type II binding sites have been subdivided into two classes, depending on their affinity for secretin (Hubel, 1972) and helodermin (Christophe et al., 1986): classical VIP binding sites exhibit low affinity for secretin (Christophe et al., 1981, 1989; Robberecht et al., 1982, 1988) whereas helodermin-preferring binding sites possess higher affinity for helodermin than for VIP or PACAP, and no affinity for secretin (Robberecht et al., 1984, 1998; Gourlet et al., 1991a;Shima et al., 1996; Solano et al., 1996). Careful characterization of125I-PACAP27 binding on membrane preparations indicated that the expression of type I and type II binding sites is not cell-specific and that most of the tissues possess various proportions of each receptor subtype (Tatsuno et al., 1990; Nguyen et al., 1993).
Pharmacological characteristics and transduction mechanisms associated with PACAP receptors
B. Biochemical Characterization of PACAP Receptors
Type I PACAP binding sites were first isolated from a tumoral cell line derived from the rat exocrine pancreas (Buscail et al., 1990). Cross-linking of 125I-PACAP27 to cell membrane preparations made it possible to isolate a 65-kDa protein (Buscail et al., 1990). In the porcine brain, type I PACAP binding sites exhibit an apparent molecular mass of 60 kDa (Schäfer and Schmidt, 1993; Schäfer et al., 1994). The extent ofN-glycosylation of type I PACAP binding sites appears to be rather low compared with other glycosylated receptors (Klueppelberg et al., 1989; Feldman et al., 1990), but it is similar to those of type II PACAP or glucagon receptors (Iwanij and Hur, 1985; Raymond and Rosenzweig, 1991). In the bovine brain, type I PACAP binding sites have a molecular mass of 57 kDa and are coupled to a Gs protein (Ohtaki et al., 1990, 1993). Type I PACAP binding sites purified from bovine brain membranes were used to sequence the N-terminal portion of the protein (Ohtaki et al., 1993). The amino acid sequence was subsequently used to clone the type I PACAP receptor (see Section III, C).
Type II PACAP binding sites have been isolated in pure form from bovine brain membranes (Ohtaki et al., 1990). The protein has an apparent molecular mass of 45 kDa, i.e., very similar to that previously reported for the VIP receptor (Couvineau et al., 1986a,b).
C. Cloning of PACAP Receptors
Three PACAP receptors have been cloned so far and termed PAC1, VPAC1, and VPAC2 receptors (Table 3) by the International Union of Pharmacology according to their relative affinity for PACAP and VIP (Harmar et al., 1998).
The PACAP-specific receptor (PAC1-R) cDNA sequence was first determined from a pancreatic acinar carcinoma cell line (Pisegna and Wank, 1993). This PAC1-R cDNA, which encodes a 495-amino acid protein with seven putative membrane-spanning domains, exhibits a high degree of sequence identity with the glucagon, secretin, and calcitonin receptor cDNAs. The PAC1-R has subsequently been cloned in humans (Ogi et al., 1993; Pisegna and Wank, 1996), bovine (Miyamoto et al., 1994), rat (Hashimoto et al., 1993; Hosoya et al., 1993; Morrow et al., 1993;Spengler et al., 1993; Svoboda et al., 1993), and mouse (Hashimoto et al., 1996b). The PAC1-R has also been cloned in the goldfishCarassius auratus (Wong et al., 1998), and the frogsRana ridibunda (Alexandre et al., 1999) and Xenopus laevis (Hu et al., 2000). Five variants resulting from alternative splicing in the third intracellular loop region have been identified in rat (Spengler et al., 1993). The splice variants are characterized by the absence (short variant) or presence of either one or two cassettes of 28 (hip or hop1 variant) or 27 (hop2 variant) amino acids (Journot et al., 1994). The presence of the hip cassette impairs adenylyl cyclase stimulation and abolishes phospholipase C (PLC) activation, suggesting that the various cassettes are involved in second messenger coupling (Table 3). In the brain and pituitary, the short variant is the most abundant form, whereas the hop variant predominates in the testes and adrenal gland (Spengler et al., 1993). A very short splice variant of PAC1-R, characterized by a 21-amino acid deletion in the N-terminal extracellular domain, has also been characterized (Pantaloni et al., 1996; Dautzenberg et al., 1999). The existence of this 21-amino acid sequence influences the receptor selectivity for the PACAP38 and PACAP27 isoforms and determines the relative potencies of the two peptides in stimulating PLC. Another PACAP receptor variant termed PAC1-R transmembrane domain (TM) 4 has been cloned in the rat cerebellum (Chatterjee et al., 1996). This latter receptor differs from the short variant of the PAC1-R by discrete sequence substitutions located in TMs II and IV. Surprisingly, activation of PAC1-R TM4 has no effect on adenylyl cyclase or PLC activity, but causes calcium influx through L-type voltage-sensitive calcium channels (Table 3). The mousePAC1-R gene spans more than 50 kb and is divided into 18 exons (Aino et al., 1995). The proximal promoter region has no apparent TATA box but contains a CCAAT box and two potential specific protein 1-binding sites that act as transcriptional activators (Dynan and Tjian, 1983; Skak and Michelsen, 1999). The rat PAC1-R gene has been localized on chromosome 4 (Cai et al., 1995) and spans 40 kb with 15 exons (Chatterjee et al., 1997). The intron/exon organization of the PAC1-R gene is very similar to that of the other members of the secretin receptor family. Alternative splicing of thePAC1-R gene also occurs in the untranslated region and could represent a regulatory mechanism involved in tissue-selective expression of the gene and/or in mRNA stability. The humanPAC1-R gene is located in region p15 of chromosome 7 (Brabet et al., 1996).
The VIP/PACAP receptor, subtype 1 (VPAC1-R) was first cloned from a rat lung cDNA library by cross-hybridization with a secretin receptor cDNA. The rat VPAC1-R cDNA encodes a 459-amino acid protein (Ishihara et al., 1992) and exhibits 50% amino acid sequence identity with the rat PAC1-R (Pisegna and Wank, 1993). The human VPAC1-R cDNA was characterized from a HT29 human colonic adenocarcinoma cell line library. The human VPAC1-R comprises 457 amino acids and possesses 84% sequence identity with the rat VPAC1-R (Sreedharan et al., 1993). The VPAC1-R gene spans 22 kb and is composed of 13 exons ranging in size from 42 to 1400 base pairs (Sreedharan et al., 1995;Pei, 1997). The promoter region encompasses several potential binding sites for nuclear factors, including specific protein 1, activator protein-2, or autotumorolytic fraction. The humanVPAC1-R gene is located on region p22 of chromosome 3 (Sreedharan et al., 1995). Selective substitution of amino acids His178 → Arg and Thr343→ Lys, Pro, or Ala by directed mutagenesis results in constitutive activation of the VPAC1-R with respect to cAMP production (Gaudin et al., 1998, 1999). The VPAC1-R also has been cloned in the goldfishCarassius auratus (Chow et al., 1997) and the frogRana ridibunda (Alexandre et al., 1999). The fact that the frog VPAC1-R exhibits pharmacological characteristics of both VPAC1 and VPAC2 receptors in mammals should help to decipher the structure-activity relationships of the VIP/PACAP receptor family.
The VIP/PACAP-receptor, subtype 2 (VPAC2-R) was cloned initially from a rat pituitary cDNA library (Lutz et al., 1993) and subsequently from a human placenta cDNA library (Adamou et al., 1995). The rat and human VPAC2-R proteins exhibit 87% amino acid identity (Gagnon et al., 1994; Svoboda et al., 1994; Adamou et al., 1995). Northern blot analysis indicates that two VPAC2-R mRNAs of 4.6 and 2.3 kb are expressed in the human skeletal muscle, heart, brain, placenta, and pancreas (Adamou et al., 1995). The VPAC2-R gene is located in region q36.3 of chromosome 7 in humans (Mackay et al., 1996), and on chromosome 4 in rats (Cai et al., 1995). The human VPAC2-R is encoded by 13 exons, and the human gene spans 117 kb (Lutz et al., 1999b).
D. Structure-Activity Relationships
A number of PACAP analogs have been synthesized to identify the determinants responsible for the recognition and activation of the receptors (Fig. 7). As previously reported for other members of the glucagon-GRF-secretin family, the N-terminal region of PACAP appears to play a crucial role for the biological activity of the peptide. In particular, it has been shown that the deletion of the His1 residue causes a 50-fold decrease in the affinity of PACAP27 for rat and human PAC1-R (Gourlet et al., 1991b; Bitar and Coy, 1993). Deletion of the His1 residue of frog PACAP38 abolishes its adenylyl cyclase-stimulating activity on adenohypophysial fragments (Yon et al., 1993b). Suppression of the His1 and Ser2 residues reduces by 3000-fold the potency of PACAP27 to stimulate adenylyl cyclase in AR4–2J rat pancreatic acinar cells (Robberecht et al., 1992a). Replacement of the Ser2 residue by Ala has little effect whereas substitution of Ser2 by Phe or Arg decreases by 1000-fold the ability of PACAP27 analogs to stimulate adenylyl cyclase (Hou et al., 1994). Substitution of the Asp3residue by Asn markedly reduces the stimulatory effect of PACAP27 on adenylyl cyclase (Hou et al., 1994). N-terminal truncated analogs of PACAP exhibit antagonistic activity of PAC1-R, indicating that the N-terminal domain is required for receptor activation but is not essential for the recognition of the binding site. Gradual deletion of the N-terminal amino acid residues of PACAP27 and PACAP38 has shown that amino acid 6 to 38 of PACAP [PACAP(6–38)] is the most potent antagonist (Robberecht et al., 1992b). Paradoxically, shorter analogs such as PACAP(14–38) retain some adenylyl cyclase-stimulating potency (Vandermeers et al., 1992). Although both PACAP27 and PACAP38 are potent agonists on PACAP/VIP receptors, the C-terminal domain appears to play a facilitatory role in the recognition of the binding sites. For instance, N-terminal truncated or substituted analogs derived from PACAP38 exhibit higher activity than their PACAP27 counterparts (Fig. 7; Vandermeers et al., 1992). The fact that a chimeric peptide formed by adding the PACAP(28–38) sequence to the VIP moiety exhibits a 100-fold higher affinity than VIP for PAC1-R (Gourlet et al., 1996a, 1997b) provides additional evidence that the C-terminal region of PACAP38 reinforces the binding efficacy of the peptide. Concurrently, the 28–38 extension may also be involved in the recognition of PACAP by specific binding proteins. In support of this notion, it has been found that one such potential binding protein, ceruloplasmin, can bind PACAP38 but not PACAP27 (Tams et al., 1999).

Structure-activity relationships of various PACAP38-related peptides. The binding affinity of a series of truncated PACAP analogs and their potency to stimulate adenylyl cyclase are indicated. Amino acid substitutions are indicated in black. a, Gourlet et al., 1991b; b, Robberecht et al., 1991b; c, Schäfer et al., 1991; d, Robberecht et al., 1992a; e, Robberecht et al., 1992b;f, Vandermeers et al., 1992; g, Ciccarrelli et al., 1994; h, Hou et al., 1994; i, Ciccarelli et al., 1995; j, Van Rampelbergh et al., 1996.
A natural peptide called maxadilan has been characterized as a selective agonist of PAC1-R (Moro and Lerner, 1997). Maxadilan is a 61-amino acid peptide that was isolated from the salivary gland of the blood-feeding sand fly Lutzomia lingipalpis on the basis of its vasodilatory activity (Lerner et al., 1991). As maxadilan does not possess any significant sequence identity with PACAP, this is a unique example of functional convergence between two peptides that do not share structural similarity. A shortened maxadilan synthetic analog, termed M65, in which the amino acid sequence 25 to 41 has been deleted, acts as a specific antagonist of PAC1-R (Uchida et al., 1998; Moro et al., 1999).
Most type II receptor antagonists designed so far are N-terminal truncated or substituted VIP peptides (Pandol et al., 1986; Turner et al., 1986; Gozes et al., 1995; Gourlet et al., 1997a). Cyclic lactam analogs of PACAP behave as potent type II receptor antagonists (Bitar et al., 1994). A cyclic peptide, RO 25–1553, acts as a selective VPAC2-R agonist with respect to binding affinity and adenylyl cyclase-stimulating potency (O'Donnell et al., 1994; Gourlet et al., 1997c). Amino acid substitutions and addition of a fatty acyl moiety have led to the development of lipophilic VIP derivatives that exhibit enhanced potency and specificity for VPAC-R (Gozes and Fridkin, 1992;Gozes et al., 1995; Gourlet et al., 1998). These data suggest that several domains are involved in the binding of PACAP to its receptors, and demonstrate the possibility of developing powerful and selective agonists or antagonists with potential therapeutic value.
The CHO and NIH 3T3 cell lines, and the yeast Saccharomyces cerevisiae, which are naturally devoid of PACAP receptors, have been widely used for the pharmacological and functional characterization of each PACAP receptor subtype after transfection (Ciccarelli et al., 1994; Delporte et al., 1995; Gaudin et al., 1996;Gourlet et al., 1996b; Van Rampelbergh et al., 1996; Hansen et al., 1999). Concurrently, the CHO and COS-7 cell lines have been used to investigate the binding properties of chimeric PACAP/VIP receptors (Vilardaga et al., 1995, 1996; Van Rampelbergh et al., 1996; Hashimoto et al., 1997; Juarranz et al., 1999b; Lutz et al., 1999a).
E. Distribution of PACAP Receptors in the CNS
The localization of PACAP binding sites and PACAP receptor mRNAs has been investigated thoroughly in the rat brain (Masuo et al., 1991,1992; Schäfer et al., 1991; Hashimoto et al., 1996a; Nomura et al., 1996; Shioda et al., 1997a; Vertongen et al., 1997b; M. Basille, D. Vaudry, Y. Coulouarn, S. Jégou, I. Lihrmann, A. Fournier, H. Vaudry and B. J. Gonzalez, submitted). The distribution and relative density of type I (PACAP-specific) and type II (PACAP/VIP) binding sites are compared in Table 4.
Localization and relative abundance of type I and type II PACAP binding sites in the rat brain
High concentrations of type I binding sites occur in various hypothalamic structures including the supraoptic nucleus (SON), the periventricular nucleus, and the lateral hypothalamic area. High densities of type I binding sites are also found in the piriform cortex, the diagonal band of Broca, the habenular nucleus, the septal nucleus, the hippocampal formation, the superficial gray layer of the superior colliculus, the dorsal raphe nucleus, and the locus ceruleus (Cauvin et al., 1991; Masuo et al., 1991, 1992; Suda et al., 1991; Hou et al., 1994). Lower concentrations of recognition sites are present in the internal granular layer of the olfactory bulb, the entorhinal cortex, the ventral posterolateral nucleus of the thalamus, the arcuate nucleus of the hypothalamus (Cauvin et al., 1991; Masuo et al., 1992;Li et al., 1997), the pineal gland (Simonneaux et al., 1998), and the granule cell layer of the cerebellum (Basille et al., 1993, 1994).
Type II binding sites are generally less abundant, and their distribution is more restricted than that of type I sites (Table 4). In the rat CNS, type II binding sites are mainly located in the olfactory bulb, the cerebral cortex, the dentate gyrus, the thalamus, and the pineal gland (Besson et al., 1984, 1986; Martin et al., 1987; Vertongen et al., 1998). In contrast, the concentration of type II binding sites is much lower than that of type I sites in many other brain regions such as the medial nucleus of the amygdaloid complex, the frontal cortex, the lateral hypothalamic nucleus, and the cerebellum (Masuo et al., 1992; Basille et al., 1993). Iodinated secretin and RO 25–1553 have been used to discriminate the respective localization of the two subclasses of type II binding sites. High concentrations of secretin-preferring sites are present in the cerebral cortex, the amygdaloid nucleus, the dentate gyrus, various thalamic nuclei, and the SON whereas RO 25–1553-preferring sites are located in the cerebral cortex, the lateral septal nucleus, the amygdaloid complex, the thalamus, the medial mammillary, periventricular, and suprachiasmatic nuclei of the hypothalamus, and the superior colliculus (Vertongen et al., 1997b). The occurrence of type I and type II binding sites on cultured astrocytes (Tatsuno et al., 1990) suggests that PACAP and/or VIP receptors are not only present on neurons but can also be expressed in glial cells (Martin et al., 1992).
The distribution and relative density of PAC1-R, VPAC1-R, and VPAC2-R mRNAs are compared in Table5. Globally, the density of PAC1-R transcript is much higher than those of the VPAC1-R and VPAC2-R transcripts (Basille et al., 2000). The expression of PAC1-R mRNA is particularly intense in the olfactory bulb, the dentate gyrus of the hippocampus, the supraoptic nuclei of the hypothalamus, the cerebellar cortex, and the area postrema (Fig. 8;Hashimoto et al., 1996a; Nomura et al., 1996; Shioda et al., 1997a;Otto et al., 1999). High levels of PAC1-R mRNA are also observed in the cingulate, entorhinal and piriform cortex, pyramidal and nonpyramidal cells of the hippocampal formation, the amygdaloid nuclei, the centromedial, mediodorsal, and ventromedial nuclei of the thalamus, the hypothalamus, the central gray, the raphe nuclei, and the superior colliculus (Hashimoto et al., 1996a; Shioda et al., 1997a). In the brain, the localization of PAC1-R transcripts correlates well with the distribution of type I binding sites (Fig.9; Basille et al., 1993; Shioda et al., 1997a). The major splice variants of PAC1-R in the rat brain is the short isoform that does not contain any hip or hop cassettes (Spengler et al., 1993; Zhou et al., 2000). The PAC1-R gene is expressed both in neurons and in glial cells (Tatsuno et al., 1991a). In neurons, PAC1-R-LI is located mainly on cell bodies and dendrites (Shioda et al., 1997a). At the ultrastructural level, accumulation of PAC1-R-immunoreactive material is observed on the plasma membrane, notably at synaptic formations (Shioda et al., 1997a). Moderate levels of PAC1-R have been detected by in situ hybridization in Bergmann glial cells in the rat cerebellar cortex (Ashur-Fabian et al., 1997). Characterization of PACAP receptor mRNA indicates that cultured glial cells express the hop2 splice variant of PAC1-R (Hashimoto et al., 1996a; Grimaldi and Cavallaro, 1999).
Localization and relative abundance of PACAP receptor mRNAs in the rat brain
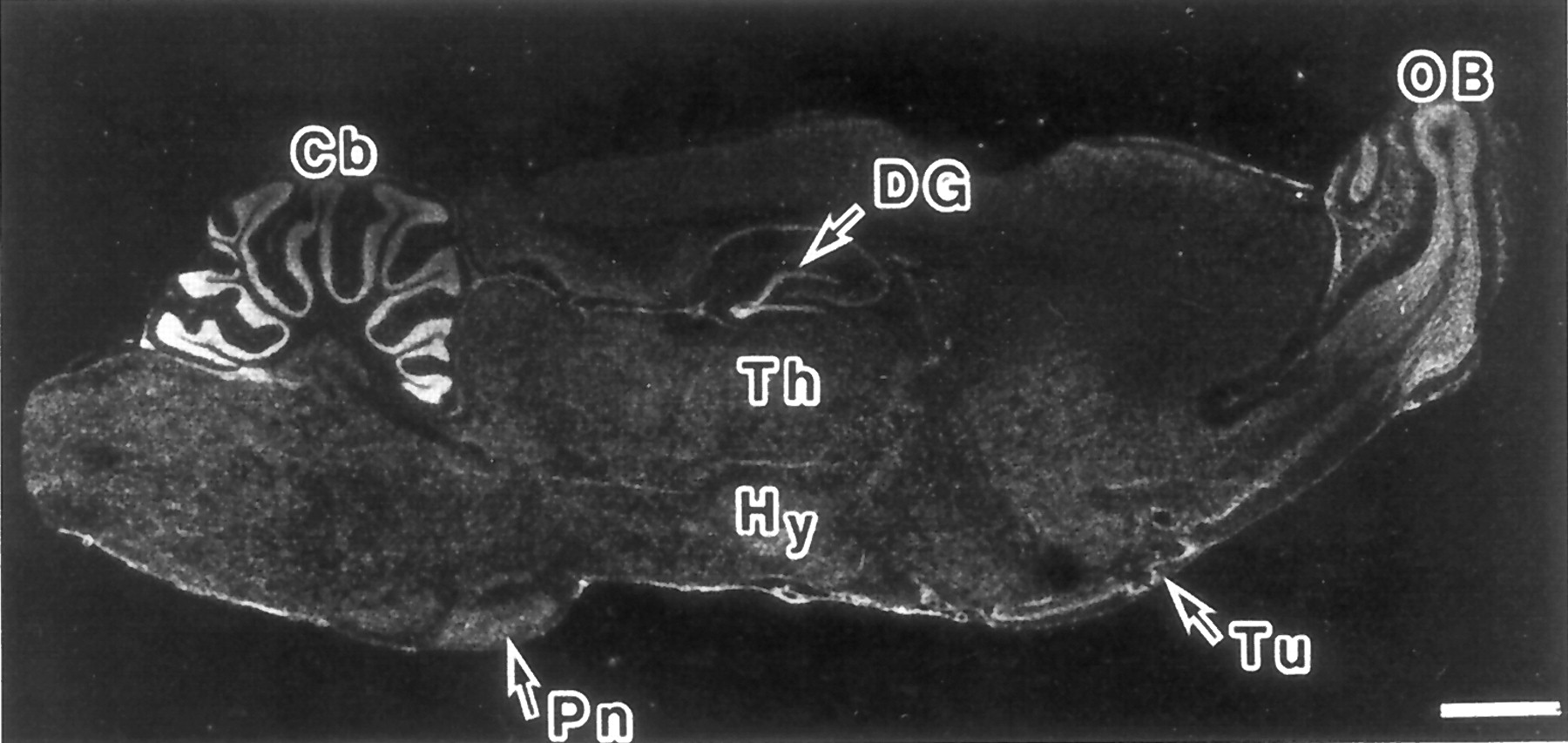
Microphotograph of a sagittal section of adult rat brain showing the pattern of expression of the PAC1 receptor mRNA as determined by in situ hybridization with a 32P-labeled cRNA probe. Cb, cerebellar cortex; DG, dentate gyrus of the hippocampus; Hy, hypothalamus; OB, olfactory bulb; Pn, pontine nuclei; Th, thalamus; Tu, olfactory tubercle. Scale bar: 2 mm. Reprinted from Shioda et al. (1997a) with permission from Neuroscience Research,Elsevier Science.
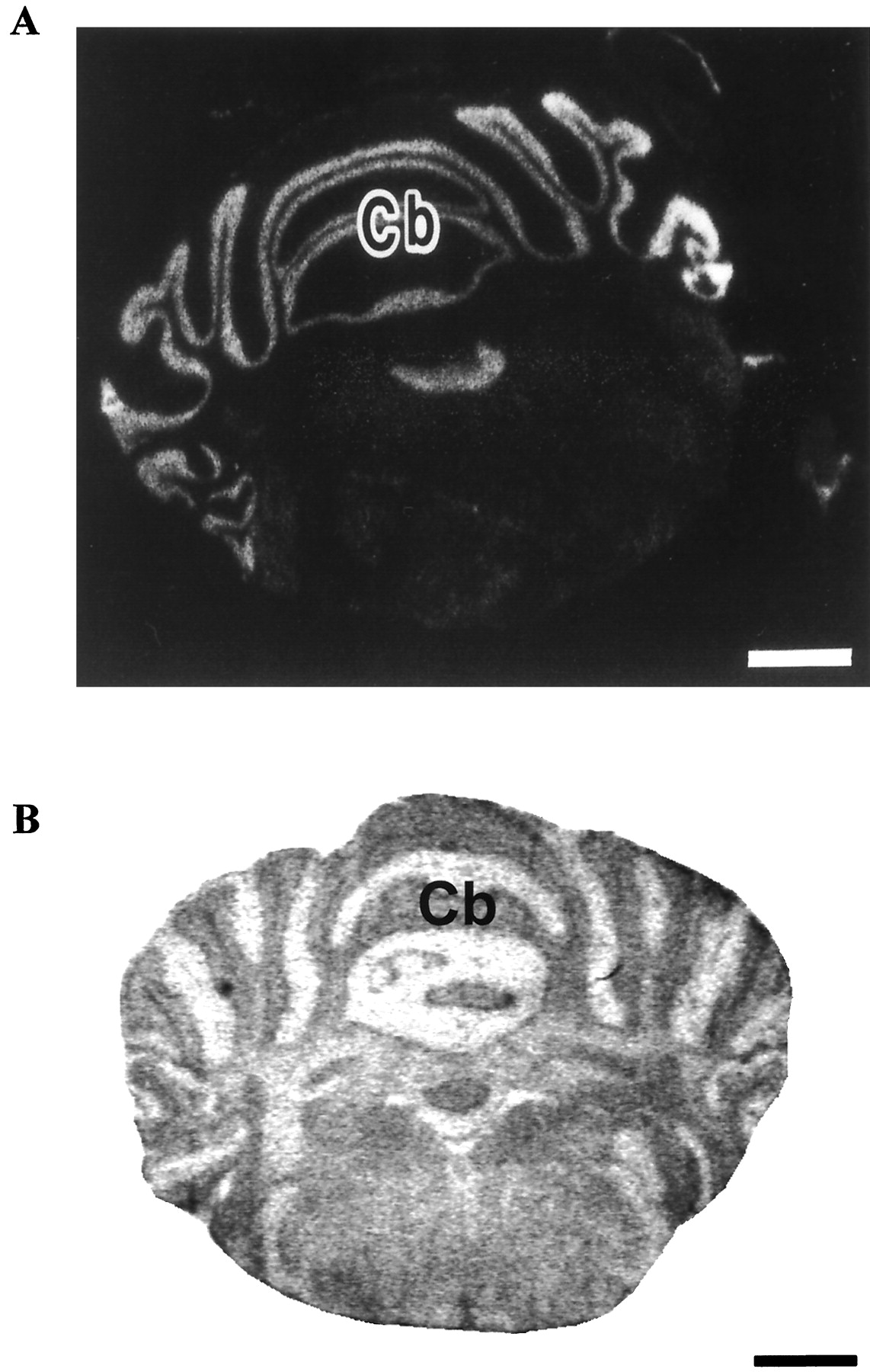
Distribution of PACAP receptors in the adult rat cerebellum. A, distribution of PAC1 receptor mRNA as determined by in situ hybridization. Reprinted from Shioda et al. (1997a) with permission from Neuroscience Research, Elsevier Science. B, expression of type I PACAP binding sites in the cerebellum. Cb, cerebellar cortex. Scale bar: 1.5 mm.
The VPAC1-R mRNA is expressed mainly in the cerebral cortex and the hippocampus (Usdin et al., 1994; Sheward et al., 1995). Anatomical mapping of the VPAC2-R mRNA demonstrates a completely different and, apparently, complementary distribution from that of the VPAC1-R mRNA (Ishihara et al., 1992; Usdin et al., 1994). Notably, a high density of VPAC2-R mRNA is present in the thalamus, the suprachiasmatic nucleus, the central nucleus of the amygdala, and the pontine nucleus (Usdin et al., 1994; Sheward et al., 1995) whereas very few VPAC1-R mRNA is found in these structures. The distribution of the VPAC2-R overlaps with that of the VPAC1-R only in the hippocampus (Usdin et al., 1994). In the olfactory bulb, VPAC1-R, and VPAC2-R, mRNAs are differentially distributed, i.e., VPAC1-R mRNA is present in the external plexiform layer whereas VPAC2-R mRNA is expressed in the internal granular layer (Usdin et al., 1994). In the cerebral cortex, VPAC1-R mRNA is abundant in layers III and V, whereas VPAC2-R mRNA is localized exclusively in layer VI. Both VPAC1-R and VPAC2-R mRNA have been characterized by reverse transcription-polymerase chain reaction on glial cells (Grimaldi and Cavallaro, 1999).
In the murine superior cervical ganglion, intense expression of PAC1-R mRNA is observed in all neurons but neither VPAC1-R nor VPAC2-R mRNAs are present (Moller et al., 1997a,b; Nogi et al., 1997b; Braas and May, 1999; DiCicco-Bloom et al., 2000). In the retina, type I PACAP binding sites predominate whereas, in the choroid, both type I and type II PACAP binding sites are expressed (Nilsson et al., 1994; D'Agata and Cavallaro, 1998). Immunocytochemical and in situ hybridization studies have revealed that PAC1-R is actively expressed in ganglion and amacrine cells as well as in the inner plexiform layer of the retina (Seki et al., 1997).
F. Distribution of PACAP Receptors in Peripheral Organs
PACAP binding sites and/or receptor mRNAs have been identified in most endocrine glands (Tables 6 and7). Type I PACAP binding sites have been characterized on rat and frog anterior pituitary membranes (Gottschall et al., 1990; Lam et al., 1990; Jeandel et al., 1999). Cytochemical labeling using biotinylated PACAP revealed that all cell types of the adenohypophysis possess PACAP recognition sites (Vigh et al., 1993;Rawlings and Hezareh, 1996). Reverse transcription-polymerase chain reaction amplification on single pituitary cells indicated that gonadotrophs express the short and hop splice variant isoforms (Bresson-Bépoldin et al., 1998). The VPAC2-R mRNA is widely distributed in the anterior pituitary whereas the VPAC1-R mRNA is not expressed (Usdin et al., 1994). In the posterior pituitary, both the neural lobe (Hashimoto et al., 1996a) and the intermediate lobe (René et al., 1996) contain moderate concentrations of PAC1-R mRNA. In the pancreas, insulin-producing cells have been shown to express both PAC1-R and VPAC2-R mRNAs (Usdin et al., 1994; Wei and Mojsov, 1996a,b; Filipsson et al., 1998a; Torii et al., 1998) whereas the VPAC1-R mRNA is found only in the walls of blood vessels (Usdin et al., 1994). In the rat adrenal gland, type I PACAP binding sites have been characterized in medullary chromaffin cells by cytoautoradiography (Shivers et al., 1991) and immunocytochemistry (Moller and Sundler, 1996). In situ hybridization studies indicate that adrenochromaffin cells actively express both the hop1 splice variant of the PAC1-R (Nogi et al., 1997a) and the VPAC1-R (Usdin et al., 1994). In contrast, the expression level of the VPAC2-R in the adrenal medulla is much lower (Usdin et al., 1994). In the frog adrenal gland, type I PACAP binding sites are expressed on both adrenocortical and chromaffin cells (Yon et al., 1994). In the rat ovary, the presence of PAC1-R and VPAC2-R mRNAs has been reported (Usdin et al., 1994; Scaldaferri et al., 1996; Kotani et al., 1997, 1998). Granulosa cells of the developing follicule express the VPAC2-R mRNA (Usdin et al., 1994) whereas the corpus luteum contains the PAC1-R mRNA (Kotani et al., 1997). In the placenta, Northern blot analysis revealed the presence of both VPAC1-R and VPAC2-R mRNA (Adamou et al., 1995; Sreedharan et al., 1995). In the testis, type I PACAP binding sites have been characterized in germ cells (Shivers et al., 1991), Leydig cells (Romanelli et al., 1997), and Sertolli cells (Heindel et al., 1992). However, identification of the mRNA by in situ hybridization indicates that the VPAC2-Rgene, but not the PAC1-R or the VPAC1-R genes, is expressed in germ cells (Usdin et al., 1994; Krempels et al., 1995;El-Gehani et al., 1998a,b). On prostate membranes, the predominant receptor subtype corresponds to the VPAC1-R (Juarranz et al., 1999a) but PAC1-R mRNA is also expressed in human benign hyperplastic prostate (Solano et al., 1999).
Localization and relative abundance of type I and type II PACAP binding sites in rat peripheral tissues
Localization and relative abundance of PACAP receptor mRNAs in rat peripheral tissues
In the digestive system, PACAP/VIP receptors are found both in the alimentary canal and accessory glands. In the human labial and submandibular gland, type II sites are found in acinar cells (Tornwall et al., 1994; Kusakabe et al., 1998). In the guinea pig stomach, type II binding sites are present in chief cells (Felley et al., 1992) whereas, in the rabbit stomach, type II sites are borne by smooth muscle cells (Murthy et al., 1997). Characterization of the receptor mRNAs confirmed that only the VPAC2-R gene is expressed in the rat, guinea pig, and rabbit stomach (Usdin et al., 1994; Teng et al., 1998). Type II binding sites are also present at different levels of the intestine (Prieto et al., 1981; Zimmerman et al., 1988, 1989). In the human colon, type II sites are located on epithelial cells (Broyart et al., 1981; Salomon et al., 1993). Type II binding sites are found on liver membranes (Guijarro et al., 1992, 1995; Gagnon et al., 1994). Characterization of the receptor mRNAs by in situ hybridization indicates that the VPAC1-R gene is predominantly expressed in the rat liver (Usdin et al., 1994).
The presence of PACAP/VIP receptors has been reported in various components of the immune system (Xin et al., 1994; Ganea, 1996). ThePAC1-R gene is expressed in rat peritoneal macrophages but not in peritoneal lymphocytes (Delgado et al., 1996a; Pozo et al., 1997). VIP-preferring sites are present in human blood mononuclear cells (Guerrero et al., 1981) and in murine splenocytes (Tatsuno et al., 1991a). The VPAC1-R gene is constitutively expressed in T-lymphocytes and thymocytes (Waschek et al., 1995a; Delgado et al., 1996c,d; Johnson et al., 1996). Stimulation through the T cell receptors-associated CD3 complex induces the expression of the VPAC2-R mRNA in T-lymphocytes (Delgado et al., 1996a).
PACAP/VIP receptors are found at all levels of the respiratory tract. In the human trachea, type II binding sites are localized in acini and excretory ducts of submucosal glands (Fischer et al., 1992). High densities of type II binding sites are also present in the lung (Lam et al., 1990; Shivers et al., 1991; Bitar and Coy, 1993; Sreedharan et al., 1995). The VPAC1-R mRNA is highly expressed in the epithelium of large bronchi whereas the VPAC2-R is present in small terminal bronchioles (Ishihara et al., 1992; Sreedharan et al., 1993; Usdin et al., 1994).
The presence of PACAP receptors has been investigated in the cardiovascular system. In the heart, the PAC1-R, VPAC1-R, and VPAC2-R have been characterized by Northern blot analysis (Gagnon et al., 1994;Adamou et al., 1995; Wei and Mojsov, 1996a,b; Wong et al., 1998). Messenger RNA encoding PAC1-R isoforms and VPAC2-R are localized in cardiac ganglia (Gagnon et al., 1994; Braas et al., 1998). The aortic tissue exhibits mRNA for all PACAP receptors (Miyata et al., 1998). However, in de-endothelized aortic tissue and cultured vascular smooth muscle cells, only VPAC2-R mRNA is detected, suggesting that VPAC2-R may mediate the vasodilator effects of PACAP (Miyata et al., 1998).
Trancripts of VPAC2-R are found in a number of other peripheral tissues such as the skeletal muscle (Wei and Mojsov, 1996a,b), the loops of Henle and the collecting tubules of the renal medulla (Usdin et al., 1994), and the white fat (Wei and Mojsov, 1996b).
G. PACAP Receptors in Tumor Cells
Neoplastic cells from breast, lung, prostate, pancreatic, colonic, and hepatocellular carcinoma often express type II PACAP/VIP binding sites (Reubi, 1995, 1999a,b; Moody et al., 1998; Busto et al., 1999). The presence of type II recognition sites has also been found in human pituitary adenoma (Robberecht et al., 1993; Oka et al., 1998) and brain glioma (Robberecht et al., 1994; Vertongen et al., 1995a). Therefore, attempts have been made currently to use iodinated VIP radioligands to localize tumor cells by scintigraphy in various tissues (Moody et al., 1998; Raderer et al., 1998; Virgolini et al., 1998;Reubi, 1999). In vitro studies have confirmed that a number of tumor cell lines express PACAP/VIP receptors. Type I binding sites have been characterized in the rat pancreatic acinar AR4–2J (Buscail et al., 1990) and medullary carcinoma 6/23 cell lines (Vertongen et al., 1994) and in the human neuroblastoma cell line NB-OK (Cauvin et al., 1990;Vertongen et al., 1997a). The hypothalamic GnRH neural cell line GT1–7 expresses the VPAC2-R gene (Olcese et al., 1997). Functional PACAP receptors have also been characterized in adrenal pheochromocytoma PC12 cells (Watanabe et al., 1990) and adrenocortical NCI-H295 cells (Haidan et al., 1998). Tumoral breast and intestinal cell lines exhibit VPAC1-R mRNA whereas neuroectodermal and pancreatic cell lines express both VPAC1-R and VPAC2-R mRNAs (Waschek et al., 1995b; Jiang et al., 1997; Madsen et al., 1998). It also has been reported that the receptor subtypes expressed in rat pituitary tumor cells are different from those found in normal adenohypophysial cells (Rawlings, 1994; Vertongen et al., 1996), suggesting a possible involvement of PACAP in the tumorigenic process.
H. Ontogenesis of PACAP Receptors
The evolution of the distribution and density of PACAP/VIP receptors has been essentially studied in the brain and adrenal gland. In the CNS, type I PACAP binding sites are detected as early as E14, and their density gradually increases throughout development to reach a plateau between 1 and 4 months (Tatsuno et al., 1994). The highest concentrations of type I PACAP binding sites are found in discrete regions of the germinative neuroepithelia at the level of the metencephalon and myelencephalon (Hill et al., 1994; Basille et al., 2000). PAC1-R mRNA is first detected in the neural tube in 9.5-day-old mouse and rat embryos (Sheward et al., 1996, 1998; Waschek et al., 1998; Zhou et al., 1999a; Jaworski and Proctor, 2000). From E9.5 to E11, the density of PAC1-R mRNA increases in the neuroepithelia of the mesencephalon and rhombencephalon (Sheward et al., 1996, 1998; Shuto et al., 1996; Zhou et al., 1999a). At E13, PAC1-R is expressed in the basal telencephalon and in the neuroepithelia of the hippocampal formation, cerebral cortex, and cerebellum (Zhou et al., 1999a). In infant rats, PAC1-R mRNA is intensively expressed in the olfactory bulb and the hippocampus (Zhou et al., 1999a). The ontogeny of type I binding sites has been investigated in detail in the rat cerebellum during postnatal development (Basille et al., 1994). In the external granule cell layer (EGL) and medulla, the density of sites is high from birth to P8, and markedly decreases from P8 to P25. In the internal granule cell layer (IGL) and molecular layer, binding sites are first detected at P8, and the density of sites gradually decreases from P8 to P25 (Basille et al., 1994). PACAP activates both adenylyl cyclase and PLC in P8 cerebellar granule cells (Basille et al., 1993, 1995;D'Agata et al., 1996). The presence of functional PACAP receptors in a germinative matrix such as the EGL suggests that PACAP may act as a trophic factor during development (See section IV, A, 4). Comparative distribution of PACAP and PACAP receptors in the developing rat brain reveals the existence of a good correlation between the localization of the peptide and its receptors in all germinative neuroepithelia, providing additional support for the involvement of PACAP as a neurotrophic factor (Masuo et al., 1994; Tatsuno et al., 1994; Sheward et al., 1996, 1998; Shuto et al., 1996; Lindholm et al., 1998; Waschek et al., 1998).
Type II PACAP binding sites are also found in the CNS at early embryonic stages, and the density of binding sites increases during postnatal development (Roth and Beinfeld, 1985). The distribution pattern of the VPAC1-R mRNA exhibits striking similarities with that of PAC1-R mRNA, although the expression level of the former is much lower than that of the latter (Pei, 1997; Basille et al., 2000). From E14 to birth, the VPAC1-R mRNA is expressed in the neuroepithelia bordering the ventricles (Pei, 1997; Basille et al., 2000). Similarly, the presence of the VPAC2-R mRNA has been detected by Northern blot analysis in the mouse brain at E14 (Waschek et al., 1996). From E21 to adulthood, the VPAC2-R mRNA is mainly detected in the suprachiasmatic nucleus of the hypothalamus and ventrolateral nucleus of the thalamus (Basille et al., 2000).
The presence of PACAP binding sites has been studied by autoradiography in the human adrenal gland during the second trimester of gestation (Yon et al., 1998). At this stage, cells derived from the ectoderm migrate inside the fetal cortical zone to form the medulla (Cooper et al., 1990; Ehrhart-Bornstein et al., 1997). In 14- to 20-week old fetuses, PACAP binding sites are exclusively located on adrenochromaffin cells (Yon et al., 1998). PACAP stimulates adenylyl cyclase activity in cultured adrenal cells, indicating that the binding sites found in the fetal human adrenal medulla actually correspond to functional receptors (Yon et al., 1998; L. Breault, L. Yon, M. Montéro, L. Chouinard, V. Contesse, C. Delarue, A. Fournier, J.G. LeHoux, H. Vaudry and N. Gallo-Payet, submitted). In newborn rats, the occurrence of PAC1-R mRNA has been reported in the medulla (Moller and Sundler, 1996). It has also been found that PACAP induces neurite outgrowth in cultured neonatal chromaffin cells (Wolf and Krieglstein, 1995). Taken together, these data suggest that PACAP may play a crucial role in the ontogenesis of the adrenal gland in mammals.
I. Phylogenetic Evolution of PACAP Receptors
The location of type I PACAP binding sites has been investigated in the CNS of the frog Rana ridibunda (Jeandel et al., 1999). The distribution pattern of PACAP binding sites appears to be very similar to that previously described in the rat brain (Shioda et al., 1997a). In particular, the olfactory bulb, pallium, striatum, habenular nuclei, and most nuclei of the thalamus contain moderate to high densities of PACAP receptors in the frog and rat (Shioda et al., 1997a; Jeandel et al., 1999). Type II PACAP binding sites have been localized in the brain of several submammalian species, including the pigeon Columba livia, the chicken Gallus domesticus, the snake Bothros atrox, the frogRana esculenta, and the fish Salmo trutta fario(Dietl et al., 1990; Hof et al., 1991; Kuenzel et al., 1997). These studies have shown that the distribution pattern of type II sites has been relatively well conserved during evolution. In particular in fish, amphibians, reptiles, and birds (Dietl et al., 1990; Hof et al., 1991) as in mammals (Martin et al., 1987; Masuo et al., 1992;Samejima et al., 1993), type II binding sites are particularly abundant in brain regions involved in the processing of specific sensory inputs.
The PAC1-R cDNA has been cloned in the goldfish Carassius auratus (Wong et al., 1998), the toad Xenopus laevis(Hu et al., 2000), the frog Rana ridibunda (Alexandre et al., 1999), and the chicken Gallus domesticus (Peeters et al., 1999). The goldfish PAC1-R exhibits 85% sequence identity with the human and rat counterparts (Wong et al., 1998). A VPAC-R cDNA has been cloned in the goldfish (Chow et al., 1997) and the frog (Alexandre et al., 1999). The frog VPAC-R cDNA exhibits the highest sequence identity (65%) with the human VPAC1-R but possesses pharmacological and tissue distribution characteristics of both mammalian VPAC1-R and VPAC2-R (Alexandre et al., 1999). Partial cDNA sequences corresponding to the spanning TMs 2 to 6 of the VPAC receptors also have been characterized in other nonmammalian species including chicken, pigeon, lizard, and salmon (Chow et al., 1997). Comparison of these partial nucleotide sequences with those of the human and rat VPAC1-R cDNAs indicates that strong evolutionary pressure has acted to conserve the primary structure of the VPAC1-R across vertebrates.
It is now well established that neuropeptide receptors frequently exist in a variety of subtypes that are encoded by distinct genes (Darlison and Richter, 1999). Because the nucleotide sequence in the protein-coding regions of the three PACAP receptor cDNAs are highly conserved (50% homology between any two receptors), it appears that the three PACAP receptor genes must have arisen from a common ancestral gene that was duplicated and subsequently diverged during the course of evolution (Ishihara et al., 1992; Lutz et al., 1993; Pisegna and Wank, 1993; Inagaki et al., 1994). The fact that the PAC1-R andVPAC2-R genes are both located on the same chromosome (human chromosome 7 and rat chromosome 4) whereas the VPAC1-R gene is located on different chromosomes (human chromosome 3 and rat chromosome 8), provides a clue regarding the evolutionary history of the three genes (Cai et al., 1995; Sreedharan et al., 1995; Brabet et al., 1996; Mackay et al., 1996). According to this observation, a first duplication would have yielded the VPAC1-R gene and a common ancestor for the PAC1-R and VPAC2-R genes. At a later stage in evolution, a second duplication of this ancestor gene would have produced two separate genes encoding PAC1-R and VPAC2-R.
IV. Biological and Pharmacological Effects of PACAP
The wide distribution of PACAP and its receptors suggests that the peptide may exert pleiotropic physiological functions. As a matter of fact, PACAP has now been shown to act as a hormone, a neurohormone, a neurotransmitter, and a trophic factor in a number of tissues.
A. Effects of PACAP on the CNS
1. Actions on the hypothalamus.
The most abundant population of PACAP-containing neurons and the highest density of PACAP binding sites are found in the hypothalamus (Tables 1 and 4; Arimura, 1992;Arimura and Shioda, 1995). In particular, a dense accumulation of PACAP-immunoreactive neurons and PACAP receptors are present in the magnocellular region of the PVN and SON where the neurosecretory perikarya producing oxytocin and vasopressin are located (Köves et al., 1990, 1991; Masuo et al., 1992; Kimura et al., 1994; Tamada et al., 1994; Hannibal et al., 1995a,b; Shioda et al., 1997b; Legradi et al., 1998). Intracerebroventricular injection of PACAP causes a marked enhancement of Fos-LI in these two hypothalamic nuclei (Nomura et al., 1999). The effects of PACAP on the electrophysiological activity of PVN and SON neurons have been studied on rat brain slices (Uchimura et al., 1996; Shibuya et al., 1998a,b). Administration of PACAP within the PVN and SON increases the firing rate activity and causes membrane depolarization of magnocellular neurons. Intracerebroventricular and intracisternal injection of PACAP causes a dose-dependent elevation of plasma vasopressin concentration (Murase et al., 1993; Seki et al., 1995b). In the neural lobe of the pituitary, PACAP stimulates the release of oxytocin and vasopressin through activation of the cAMP/protein kinase A (PKA) signaling pathway (Lutz-Bucher et al., 1996).
PACAP has also been shown to modulate the activity of various other hypothalamic neuronal populations. For instance, central administration of PACAP produces significant increases in GnRH, somatostatin, and CRF gene expression, which are prevented by concomitant injection of the PACAP antagonist PACAP(6–38) (Li et al., 1996; Grinevich et al., 1997). Intracerebroventricular injection of PACAP enhances the level of the dopamine metabolite DOPAC in the sheep medial basal hypothalamus (Anderson and Curlewis, 1998) and stimulates the expression of PRL mRNA in the rat hypothalamus (Bredow et al., 1994). In the ovariectomized ewe, infusion of PACAP in the arcuate nucleus of the hypothalamus reduces plasma PRL concentration (Anderson et al., 1996). Similarly, injection of PACAP in the medial basal hypothalamus suppresses luteinizing hormone (LH) secretion and LH pulse frequency (Anderson et al., 1996). Taken together, these data indicate that PACAP may act within the hypothalamus as a neurotransmitter or neuromodulator to regulate the secretion of neurohypophysial and hypophysiotropic neurohormones.
In rat, daily variations in the density of PAC1-R mRNA are observed in the suprachiasmatic and supraoptic nuclei with two peaks at noon and midnight, but not in the cingulate cortex (Cagampang et al., 1998). Similar biphasic variations of VPAC2-R mRNA levels are observed in the suprachiasmatic nucleus (Cagampang et al., 1998; Shinohara et al., 1999). These results indicate that PACAP receptors are differentially expressed in the rat brain across the 24-h cycle, suggesting that PACAP is involved in the circadian pacemaker clock. Consonant with this hypothesis, injection of PACAP at the vicinity of the suprachiasmatic nucleus has been found to reset the circadian clock in a manner similar to light (Chen et al., 1999a; Gillette and Tischkau, 1999;Harrington et al., 1999).
2. Actions of PACAP on the pineal gland.
Circadian variations in PACAP content occur in the rat pineal (Fukuhara et al., 1998), and a high density of PACAP binding sites is present in the pineal gland (Table 4; Masuo et al., 1992; Simonneaux et al., 1998), suggesting that PACAP is involved in the regulation of the rhythmicity of melatonin production. Exposure of pinealocytes to graded concentrations of PACAP causes a dose-dependent increase in the activity of two key enzymes of the melatonin biosynthetic pathway, serotonin-N-acetyltransferase (Yuwiler et al., 1995) and hydroxyindole-O-methyltransferase (Ribelayga et al., 1997). Consistent with these observations, PACAP has been found to stimulate melatonin secretion by perifused rat pineal gland (Simonneaux et al., 1993) and cultured pinealocytes (Chik and Ho, 1995; Simonneaux et al., 1998). The stimulatory action of PACAP on melatonin release is associated with calcium influx through L-type calcium channels (Chik et al., 1997) and phosphorylation of cAMP-responsive element-binding protein (CREB) (Schomerus et al., 1996, 1999). The effect of PACAP on CREB phosphorylation culminates in the first part of the dark period of the 24-h cycle (Maronde et al., 1997) in concomitance with the peak of PACAP content in the pineal gland (Fukuhara et al., 1998). PACAP causes phosphorylation of CREB in the suprachiasmatic nucleus during the light period, and the effect of PACAP on CREB phosphorylation is suppressed by melatonin (Vanecek et al., 1987; Kopp et al., 1997; Von Gall et al., 1998). Similarly, melatonin suppresses the PACAP-induced stimulation of cAMP production in the whole chicken hypothalamus and in the rat suprachiasmatic nucleus and pituitary cells, indicating that the hypothalamus is a site for a functional interaction between PACAP and the pineal hormone melatonin (Von Gall et al., 1998; Kopp et al., 1999;Nowak et al., 1999; Slamar et al., 2000).
3. Behavioral actions.
A number of neuropeptides have been shown to participate in the control of appetite and feeding behavior (Kalra et al., 1999). In particular, neuropeptide tyrosine (NPY), which, contrary to PACAP, inhibits adenylyl cyclase activity (Chance et al., 1989), is a highly potent orexigenic peptide (Clark et al., 1984; Zimanyi et al., 1998). The high concentration of PACAP-expressing neurons in the PVN and ventromedial hypothalamic nuclei (Table 1; Arimura, 1992; Arimura and Shioda, 1995; Hannibal et al., 1995a; Legradi et al., 1998), two hypothalamic regions that play a role in the regulation of food intake (Luiten et al., 1987; Leibowitz, 1988), suggests that PACAP could be involved in the control of food consumption (Christophe, 1998). Indeed, i.c.v. injection of PACAP decreases food uptake (Morley et al., 1992; Chance et al., 1995; Mizuno et al., 1998) and antagonizes the orexigenic effect of NPY (Morley et al., 1992). Concurrently, injection of PACAP in the vicinity of the perifornical lateral hypothalamus stimulates drinking (Puig de Parada et al., 1995); reciprocally, water deprivation causes an increase in PACAP-LI in cell bodies and nerve fibers of the subfornical organ (Nomura et al., 1997), suggesting that PACAP may play a role in the regulation of dipsic behavior.
Intracerebroventricular injection of PACAP enhances grooming (Morley et al., 1992) and increases the motor activity and the rearing behavior in rat (Masuo et al., 1995). Central administration of PACAP or VIP at the onset of darkness enhances rapid eye movement sleep (Bredow et al., 1994; Fang et al., 1995; Bourgin et al., 1997; Ahnaou et al., 1999). Intrathecal injection of PACAP suppresses the flexion reflex induced by electrical stimulation of the plantar nerve (Zhang et al., 1993a). The possible effect of PACAP in the transmission of noxious stimuli is currently a matter of debate: PACAP has been found to reduce the instances of flinching behavior in the formalin test, indicating that the peptide may possess antinociceptive properties (Yamamoto and Tatsuno, 1995), whereas other reports suggest that PACAP may play a facilitatory role in pain transmission (Narita et al., 1996; Xu and Wiesenfeld-Hallin, 1996; Dickinson et al., 1997,1999; Dickinson and Fleetwood-Walker, 1999; Mulder et al., 1999). The fact that the Drosophila memory gene amnesiac encodes a peptide with significant sequence similarity with PACAP indicates that, in invertebrates as in vertebrates, PACAP and related peptides could also exert behavioral activities (DeZazzo et al., 1999).
4. Neurotrophic actions.
The presence of high concentrations of PACAP and PACAP receptors in germinative areas of the developing brain indicates that the peptide may exert important functions during ontogenesis of the CNS. Indeed, in cerebellar granule cells cultured in conditions promoting apoptosis, PACAP inhibits programmed cell death (Fig. 10; Cavallaro et al., 1996; Chang et al., 1996; Campard et al., 1997; Gonzalez et al., 1997a; Villalba et al., 1997; Vaudry et al., 2000) and stimulates neurite outgrowth (Fig.10; Gonzalez et al., 1997a). Second messenger studies have been conducted to investigate the mechanisms involved in the neurotrophic activity of PACAP (Fig. 11; Gonzalez et al., 1997b). Activation of PAC1-R induces a dose-dependent stimulation of cAMP production and polyphosphoinositide hydrolysis (Gonzalez et al., 1994; Basille et al., 1995; Favit et al., 1995; Villalba et al., 1997). In vitro experiments have shown that the effect of PACAP on cell survival is mediated through activation of the adenylyl cyclase pathway, leading to phosphorylation of the extracellular signal-regulated (ERK)-type of mitogen-activated protein (MAP) kinase (Villalba et al., 1997) and to an increase in c-fos gene expression (Fig. 11; Vaudry et al., 1998a,b). In cultured granule cells, PACAP also stimulates calcium mobilization (Gonzalez et al., 1996; Mei, 1999) and blocks transient potassium currents (Zerr and Feltz, 1994), two processes often involved in programmed cell death regulation (Colom et al., 1998; Kobayashi and Mori, 1998; Krebs, 1998). The effect of PACAP on the development of the rat cerebellum has been investigated recently in vivo (Vaudry et al., 1999). Injection of PACAP at the surface of the cerebellum of 8-day-old pups induces a transient enlargement of the volume of the cerebellar cortex (Fig.12), with a maximun effect at P12, which can be accounted for by an increase in the number of granule cells in the EGL, the molecular layer, and the IGL (Fig.13). The effect of PACAP on the number of granule cells is blocked by the antagonist PACAP(6–38). The fact that the PACAP antagonist produces by itself a slight inhibition of the number of granule cells in the IGL indicates that endogenous PACAP may exert a physiological role in the development of the rat cerebellum (D. Vaudry, B. J. Gonzalez, M. Basille, T. P. Pamantung, A. Fournier and H. Vaudry, submitted).

Microphotographs illustrating the effect of PACAP38 on cell survival and neurite outgrowth of rat cerebellar granule cells after 48 h of culture. Scale bar: 25 μm. Reprinted from Gonzalez et al. (1997) with permission fromNeuroscience, Elsevier Science.
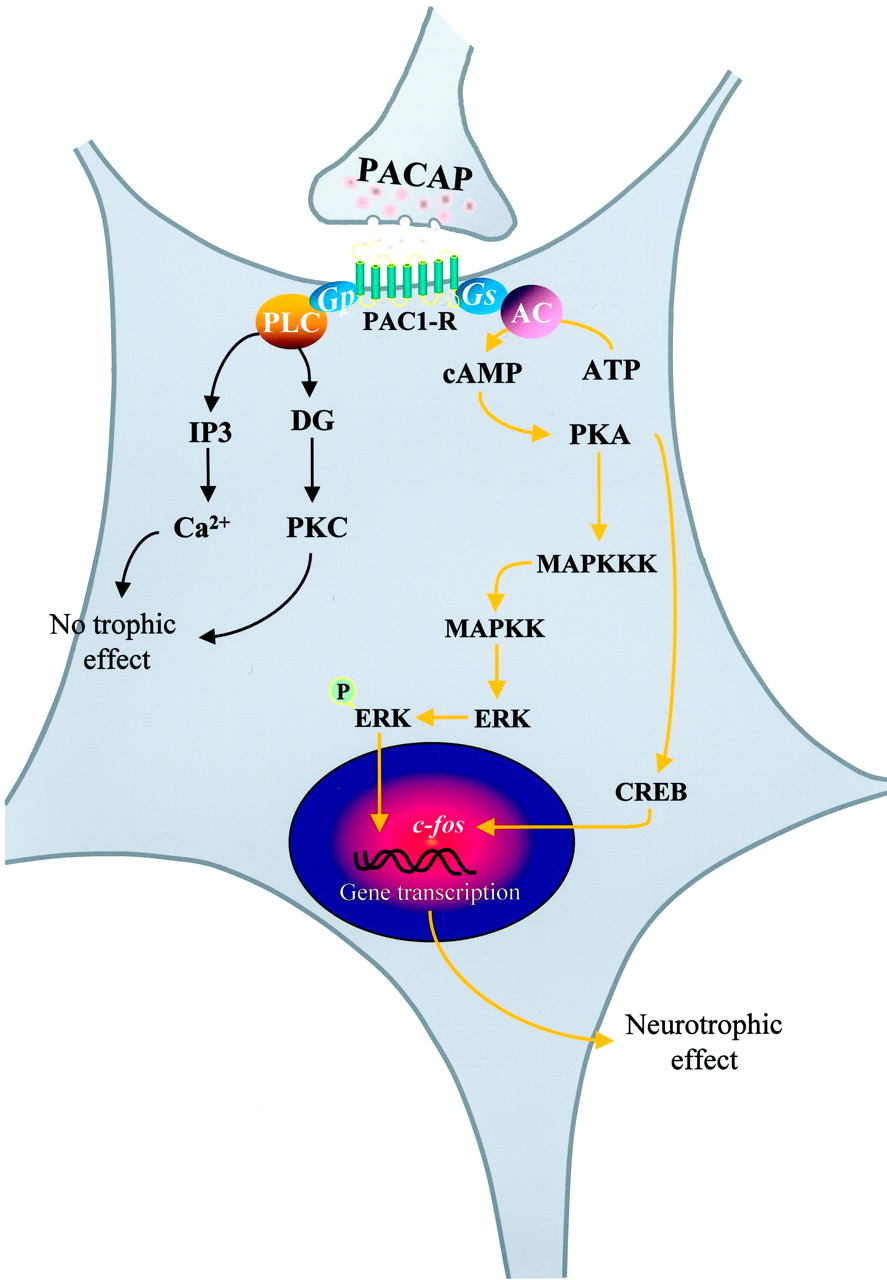
Schematic representation of the intracellular mechanisms likely involved in the trophic activity of PACAP on cerebellar granule cells. AC, adenylyl cyclase; DG, diacylglycerol; IP3, inositol trisphosphate; MAPKK, mitogen-associated protein-kinase-kinase; MAPKKK, MAPKK-kinase. (Adapted from Vaudry et al., 1998a,b).
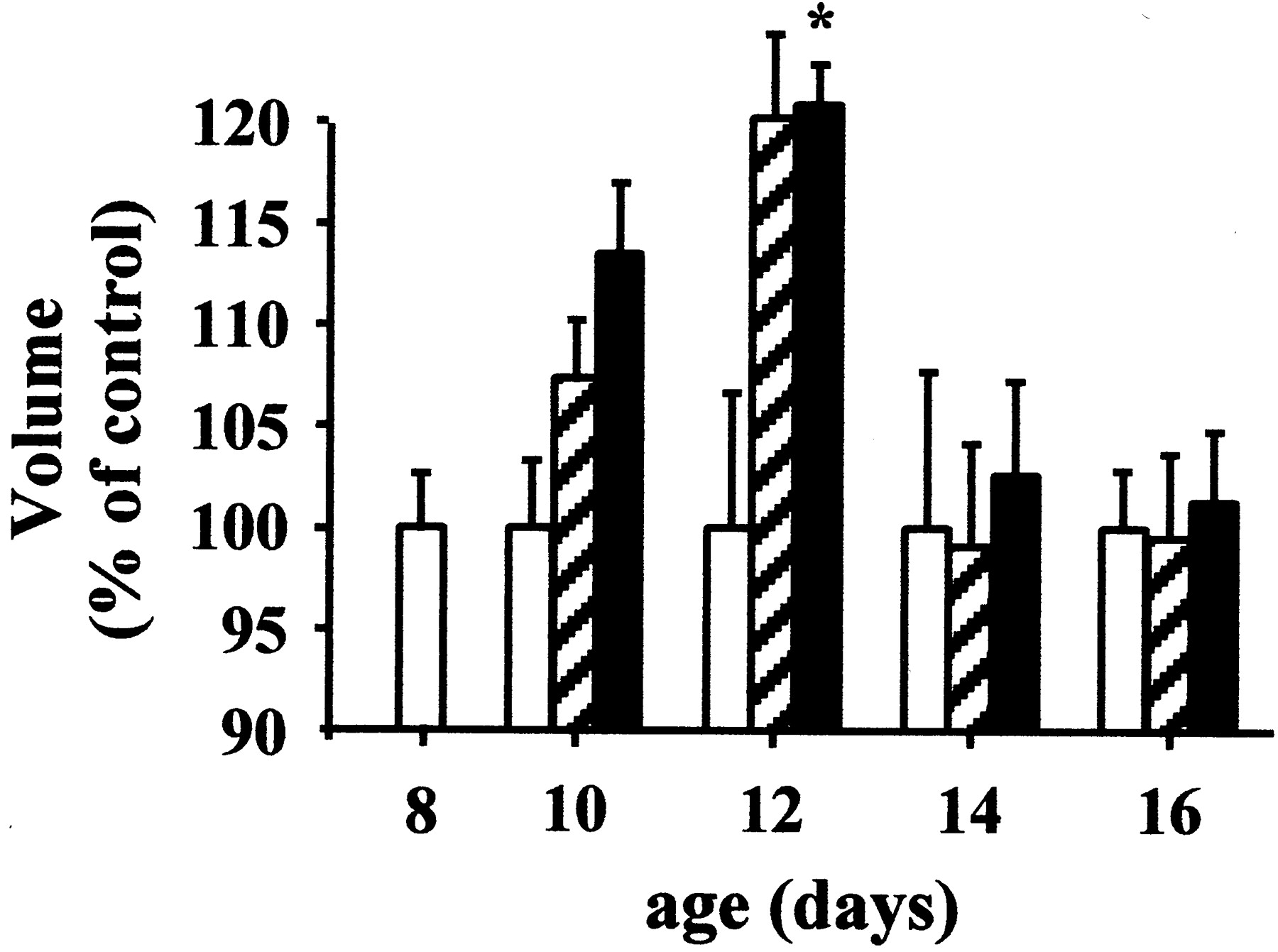
Time course of the effect of PACAP on the volume of the cerebellar cortex. Eight-day-old (P8) rats were treated with saline (open columns), 0.01 μg of PACAP38 (hatched columns), or 1 μg of PACAP38 (filled columns) up to P16. *P < .05 versus control. Reprinted from Vaudry et al. (1999) with permission from the Proceedings of the National Academy of Sciences of the USA, the National Academy of Sciences of the USA.

Effects of PACAP on the histogenesis of the cerebellar cortex. Eight-day-old (P8) rats were injected with saline (control) or 1 μg PACAP38 at the surface of the cerebellar cortex, and the thickness of the EGL, molecular layer (Mol), and IGL were measured at P12. *P < .05 as compared with control. Scale bar: 100 μm. Reprinted from Vaudry et al. (1999) with permission from the Proceedings of the National Academy of Sciences of the USA, the National Academy of Sciences of the USA.
On cortical neuron precursors, PACAP decreases the proportion of mitotic cells and promotes neuroblast differentiation, indicating that the peptide is also involved in the development of the cerebral cortex (Lu and DiCicco-Bloom, 1997; Lu et al., 1998; DiCicco-Bloom et al., 1998). After a week of culture in the presence of serum, cortical neuroblasts turn into mature neurons that express glutamate and its receptors. It should be recalled that micromolar concentrations of glutamate exert a slight protective action on cortical neurons in primary culture whereas millimolar doses of glutamate induce apoptotic cell death (Choi et al., 1987; Koh et al., 1991; Sagara and Schubert, 1998). In this model, PACAP potentiates the effect of otherwise marginally effective concentrations of glutamate (≈1 μM) onc-fos expression (Martin et al., 1995), arachidonic acid release (Stella and Magistretti, 1996; Magistretti et al., 1998a), and brain-derived neurotrophic factor production (Pellegri et al., 1998). In contrast, PACAP protects cultured cortical neurons from the cytotoxic effects of high (≈1 mM) concentrations of glutamate (Morio et al., 1996). A neuroprotective effect of PACAP on glutamate-induced neurotoxicity also has been reported in cultured retinal neurons (Shoge et al., 1999). Most of the actions of PACAP on cortical neurons are mediated through the cAMP pathway (Martin et al., 1995; Morio et al., 1996) although it has been reported that PACAP can directly modulate N-methyl-d-aspartate receptors independently of intracellular second messengers (Liu and Madsen, 1997, 1998). On these neurons, PACAP prevents the neurotoxic effect of lipopolysaccharide administration (Kong et al., 1999). In mesencephalic dopaminergic neurons, PACAP counteracts the effects of 6-hydroxydopamine neurotoxicity (Takei et al., 1998). Ischemic death of hippocampal neurons can be prevented by infusing PACAP (Uchida et al., 1996). PACAP is still effective to protect cell death when treatment is started 24 h after the ischemia, which suggests that PACAP may have therapeutic potency in treating cerebral injuries. After focal cerebral ischemia, the tumor suppressor p53 and the zinc finger protein Zac-1 (two genes responsible for cell cycle arrest and apoptosis control) are up-regulated (Gillardon et al., 1998;Ciani et al., 1999). The p53 and Zac proteins have been demonstrated to regulate the PAC1-R gene, which could, in the presence of its agonist, counteract the damages of ischemia. Consistent with this finding, PACAP and PAC1-R mRNA have been shown to increase in the cortex and the hippocampus after traumatic brain injury (Skoglösa et al., 1999a). Taken together, these studies suggest that during development, PACAP acts as a neurotrophic factor whereas, in the adult brain, the peptide appears to function as a neuroprotective agent that attenuates the neuronal damage resulting from various insults (Arimura, 1998; Brenneman et al., 1999). A 4-amino acid lipophilic fragment of PACAP (stearyl-Lys-Lys-Tyr-Leu-NH2) that offers enhanced bioavailability and stability has been developed, and it has been reported that intranasal administration of this PACAP derivative provides neuroprotection in vivo (Gozes et al., 1999).
In the dorsal root ganglions of embryos and newborn rats, thePACAP gene is expressed in sensory neurons (Lioudyno et al., 1998), and the levels of PACAP and PAC1-R mRNAs are up-regulated by axotomy (Zhang et al., 1996, 1998; Zhou et al., 1999b). Treatment of cultured ganglion neurons with PACAP increases cell survival and promotes neurite outgrowth (Lioudyno et al., 1998), supporting the view that PACAP exerts beneficial effects in nerve restoration after injury.
5. Actions on glial cells.
Consistent with the occurrence of PACAP receptors in astroglial cells, PACAP has been shown to mobilize intracellular calcium stores (Tatsuno and Arimura, 1994) and to activate a quinine-sensitive potassium outward current (Ichinose et al., 1998) in rat astrocytes. In brain slices from newborn rat, PACAP enhances the number of glial precursor cells that express the proenkephalin-A gene in the neocortical subventricular zone of the rat brain (Just et al., 1998). In cultured astrocytes, PACAP also stimulates the MAP kinase ERK2, suggesting that PACAP may regulate proliferation of astroglial cells (Moroo et al., 1998). In support of this notion, in vivo administration of a VIP antagonist induces a dramatic reduction of the density of astrocytes in the cortex of E17 mouse embryos, and this effect is reversed by cotreatment with PACAP or the VPAC2-R agonist RO 25–1553 (Zupan et al., 1998), indicating that PACAP is actually involved in neocortical astrocytogenesis. In astrocytes, PACAP increases the production of neurotrophic factors that are responsible for neuronal proliferation and/or differentiation (Ashur-Fabian et al., 1997). In this respect, PACAP has been shown to reduce ciliary neurotrophic factor mRNA level (Nagao et al., 1995). In contrast, PACAP activates brain-derived neurotrophic factor, a trophic peptide involved in neuronal plasticity (Pellegri et al., 1998) and stimulates the secretion of interleukin (IL)-6, which acts as a trophic cytokine in the CNS (Gottschall et al., 1994). Surprisingly, some of the neuroprotective effects of VIP that involve astrocytes cannot be mimicked by PACAP, suggesting the involvement of a VIP-specific receptor that remains to be characterized (Gressens et al., 1997,1998a,b, 1999; Hill et al., 1999).
B. Effects of PACAP on the Pituitary Gland
The ability of PACAP to stimulate cAMP formation in pituitary cells provided the first evidence that the peptide may act as a hypophysiotropic neurohormone (Christophe, 1993; Arimura and Shioda, 1995; Nussdorfer and Malendowicz, 1998). The action of PACAP on the adenohypophysis has been reviewed in detail by Rawlings and Hezareh (1996). Among the different hypophysiotropic neuropeptides identified so far, the situation of PACAP is rather unique in that PACAP receptors are expressed by all endocrine cell types and by folliculo-stellate (FS) cells of the adenohypophysis (Vigh et al., 1993). Cytofluorometric studies, conducted on dispersed rat pituitary cells, have shown that PACAP actually induces calcium mobilization in all categories of endocrine cells (Canny et al., 1992; Gracia-Navarro et al., 1992;Rawlings et al., 1993, 1994; Hezareh et al., 1996; Rawlings and Hezareh, 1996). Consistent with this observation, PACAP stimulates the release of GH, adrenocorticotropic hormone (ACTH), LH, follicle-stimulating hormone (FSH), and PRL (Goth et al., 1992; Hart et al., 1992; Leonhardt et al., 1992; Coleman and Bancroft, 1993; Arbogast and Voogt, 1994; Coleman et al., 1996; Koch and Lutz-Bucher, 1993;Perrin et al., 1993; Hashizume et al., 1994; Velkeniers et al., 1994;Martinez-Fuentes et al., 1998c; Ortmann et al., 1999). The effects of PACAP on the different pituitary cell types are summarized in Table8.
Effects of PACAP on pituitary cells
Gonadotrope cells.
Gonadotropin secretion is predominantly regulated by GnRH (Conn et al., 1981; Waters and Conn, 1991). There is now evidence that PACAP acts either alone or synergistically with GnRH to stimulate LH and FSH mRNA expression (Tsujii et al., 1995; Tsujii and Winters, 1995; McArdle and Counis, 1996; Winters et al., 1997) and gonadotropin secretion (Culler and Paschall, 1991; Schomerus et al., 1994; Tsujii et al., 1994; Tsujii and Winters, 1995; Petersen et al., 1996; Ortmann et al., 1999). In the male rat, intra-atrial injection of PACAP, but not VIP, increases plasma LH level (Leonhardt et al., 1992;Osuga et al., 1992). Perifusion of rat anterior pituitary cells with PACAP induces a transient stimulation of gonadotropin release and a concomitant increase in cytosolic calcium concentration (Canny et al., 1992; Rawlings et al., 1994; Tsujii et al., 1994). The effect of PACAP on gonadotropin mRNA expression involves the cAMP/PKA pathway (Ishizaka et al., 1993; Winters et al., 1997) whereas the stimulatory effect on FSH/LH release is under the control of calcium mobilization (Canny et al., 1992; Masumoto et al., 1995).
Somatotrope cells.
Secretion of GH is stimulated by GRF and inhibited by somatostatin (Sheppard et al., 1985). Administration of PACAP to cultured pituitary cells causes a significant increase in both GH mRNA expression and GH release (Velkeniers et al., 1994; Rousseau et al., 1999). In contrast, PACAP does not modify GH secretion from superfused cells (Velkeniers et al., 1994), suggesting that the stimulatory effect of PACAP on pituitary cells in static incubation involves the paracrine mediation of other hormones. PACAP exerts an additive effect on GRF-stimulated GH output (Hashizume et al., 1994), and the stimulatory activity of PACAP on GH release is inhibited by the addition of somatostatin (Goth et al., 1992; Hashizume et al., 1994). PACAP increases intracellular calcium concentration in frog and rat somatotrope cells (Canny et al., 1992; Gracia-Navarro et al., 1992;Yada et al., 1993), and the PACAP-evoked calcium response is blocked by the PKA antagonist Rpc-AMPs, indicating that the effect of PACAP is mediated through activation of the cAMP/PKA pathway (Rawlings et al., 1993, 1995). The elevation of cytosolic calcium plays a pivotal role in PACAP-induced GH secretion (Martinez-Fuentes et al., 1998a,b,c). The maximal effect of PACAP on GH release is observed after 15 min of treatment whereas prolonged incubation or pretreatment with PACAP causes desensitization of the secretory response (Goth et al., 1992;Wei et al., 1993).
Lactotrope cells.
The secretion of PRL is predominantly under the tonic inhibitory control exerted by dopamine (Martinez de la Escalera and Weiner, 1992). The secretory activity of lactotrope cells is also regulated by various hypothalamic neuropeptides (Ruberg et al., 1981; Carbajal and Vitale, 1997). In particular, VIP and to a lesser extent PHI and secretin stimulate PRL secretion (Vijayan et al., 1979;Enjalbert et al., 1980; Kimura et al., 1987; Muratori et al., 1994;Judd, 1995; Youngren et al., 1998). It also has been shown that VIP and PHI enhance the electrophysiological activity of lactotrope cells (Hedlund et al., 1988). In fact, VIP is synthesized (Arnaout et al., 1986) and released by rat lactotrope cells (Nagy et al., 1988), indicating that VIP and PHI could act as autocrine stimulators of PRL secretion. The observation that VIP-related peptides stimulate lactotrope cell activity prompted several groups to investigate the ability of PACAP to modulate PRL secretion. Intravenous injection of PACAP to anesthetized rats induces a 4-fold increase of plasma PRL concentration (Leonhardt et al., 1992; Yamauchi et al., 1995). The effect of systemic administration of PACAP can be accounted for, at least in part, by a direct action at the pituitary level because the peptide can also enhance plasma PRL level in hypothalamus-lesioned animals (Jarry et al., 1992). In vitro studies have confirmed that PACAP exerts a direct stimulatory effect on cytosolic calcium concentrations in frog and rat lactotrope cells (Canny et al., 1992;Gracia-Navarro et al., 1992). However, it should be noted that PACAP increases the intracellular calcium level in 45% of PRL cells in frog (Gracia-Navarro et al., 1992) but only in 9% of PRL cells in rat (Canny et al., 1992). Studies aimed at investigating the effect of PACAP on PRL secretion by pituitary cells have led to controversial results. It has been initially reported that PACAP is devoid of PRL-releasing activity in cultured rat adenohypophysial cells (Miyata et al., 1989). PACAP was also found to have no effect on PRL release from cultured ovine (Sawangjaroen et al., 1997) and bovine (Hashizume et al., 1994) pituitary cells. In contrast, other studies have shown that PACAP can either stimulate (Nagy et al., 1993) or inhibit (Jarry et al., 1992) PRL release from rat pituitary cells. To elucidate the origin of these apparent discrepancies, the effects of PACAP on PRL secretion have been compared in cultures of dispersed or aggregated cells and in pituitary fragments (Benter et al., 1995). In monolayer cultures, PRL release was inhibited by PACAP, whereas in cultures of aggregated cells and in pituitary fragments, PRL output was stimulated (Benter et al., 1995). These data suggest that cell-to-cell communication plays a crucial role in determining the type of action of PACAP on PRL secretion. Whereas i.v. injection of PACAP produces a significant increase in plasma PRL concentration in rat (Jarry et al., 1992; Leonhardt et al., 1992; Yamauchi et al., 1995), systemic administration of PACAP has no effect on PRL level in sheep (Sawangjaroen and Curlewis, 1994), suggesting the existence of marked species differences.
Besides its hypophysiotropic action at the pituitary level, PACAP may also regulate PRL secretion through modulation of various hypothalamic factors. In particular, in sheep, injection of PACAP into the medial basal hypothalamus stimulates dopamine release from tuberoinfundibular neurons leading to an inhibition of PRL secretion (Anderson and Curlewis, 1998). PACAP has also been found to decrease the activity of pyroglutamyl peptidase II (Vargas et al., 1998), a TRH-specific ectoenzyme that cleaves the pyroglutamyl-histidyl peptide bond of TRH (Charli et al., 1998). Because TRH is known to activate PRL secretion (Martinez de la Escalera and Weiner, 1992), the inhibition of pyroglutamyl peptidase II induced by PACAP may cause indirect stimulation of PRL release through reduction of TRH degradation. Taken together, these data indicate that PACAP may affect PRL secretion either via presynaptic action on hypothalamic neurons or via postsynaptic regulation of the activity of hypophysiotropic neurohormones.
Corticotrope cells.
The secretion of ACTH is primarily regulated by CRF (Rivier et al., 1982a) and PACAP has been shown to activate CRF gene expression in the rat PVN (Grinevich et al., 1997). Intravenous administration of PACAP provokes a dose-related increase in plasma ACTH level in rat (Leonhardt et al., 1992) and human (Chiodera et al., 1996). The effect of PACAP on circulating ACTH in human is not mimicked by VIP, indicating that the peptide acts through PAC1-R. In vitro, PACAP stimulates ACTH secretion from superfused (Miyata et al., 1989) or cultured rat pitutary cells (Hart et al., 1992). However, in rat, the effect of PACAP on ACTH secretion by cultured cells does not reach significance until 24 h, suggesting that PACAP does not exert a direct stimulatory action on corticotrope cells (Hart et al., 1992). Other in vitro studies have shown that PACAP does not stimulate ACTH secretion from rat pituitary cells within 3 h of incubation (Culler and Paschall, 1991; Koch and Lutz-Bucher, 1993). In the frogRana rididunda, PACAP causes an increase in cytosolic calcium concentration in 25% of corticotrope cells (Gracia-Navarro et al., 1992) and stimulates ACTH secretion within 4 h (Martinez-Fuentes et al., 1994), indicating that, in amphibians, PACAP directly activates corticotrope cells.
Thyrotrope cells.
In vivo administration of PACAP does not affect plasma thyroid-stimulating hormone (TSH) concentrations in rat (Hart et al., 1992) and human (Chiodera et al., 1996). Consistent with this observation, PACAP does not modify TSH secretion from cultured rat anterior pituitary cells (Culler and Paschall, 1991), and only a few thyrotrope cells express PACAP binding sites (Vigh et al., 1993). In frog, PACAP increases free cytosolic calcium concentration in thyrotrope cells (Gracia-Navarro et al., 1992) but has no effect on TSH release (Martinez-Fuentes et al., 1994).
FS cells.
Besides endocrine cells, the anterior pituitary encompasses a population of glial-like cells named FS cells. Incubation of cultured rat FS cells with PACAP causes stimulation of cAMP formation and IL-6 production (Tatsuno et al., 1991c). Similarly, PACAP increases cAMP level and stimulates vascular endothelial growth factor and IL-6 secretion in the mouse FS-like cell line TtT/GF (Matsumoto et al., 1993; Gloddek et al., 1999). Because IL-6 is involved in the differentiation of pituitary cells (Renner et al., 1998) and stimulates the release of various adenohypophysial hormones (Renner et al., 1998), several indirect effects of PACAP on endocrine pituitary cells may be mediated through activation of FS cells (Benter et al., 1995). In support of this notion, FS cells have been shown to play a pivotal role in paracrine interactions within the anterior pituitary (Baes et al., 1987; Allaerts and Denef, 1989; Valentijn et al., 1998).
Pituitary fibroblasts.
The anterior pituitary gland also contains fibroblasts, a type of agranular connective cells (Gospodarowicz, 1979). PACAP has been shown to stimulate cAMP formation in cultured pituitary fibroblasts, suggesting that the peptide may modulate fibroblast proliferation (Koch and Lutz-Bucher, 1992b).
Melanotrope cells.
The intermediate lobe of the pituitary is composed of a homogeneous population of cells, the melanotrope cells, which express the multifunctional precursor protein proopiomelanocortin (POMC) (Crine et al., 1978). Post-translational processing of POMC in melanotrope cells gives rise to the formation of the melanotropin α-melanocyte-stimulating hormone (α-MSH) and the opioid peptide β-endorphin (Mains and Eipper, 1979). In rat, PACAP stimulates cAMP production and α-MSH release in cultured melanotrope cells (Koch and Lutz-Bucher, 1992a). PACAP has also been found to increase POMC mRNA level in the rat pars intermedia (René et al., 1996). The stimulatory effect of PACAP on POMC gene expression and α-MSH secretion is associated with calcium influx throughl-type calcium channels (Tanaka et al., 1997b). The occurrence of PACAP mRNA in the neurointermediate lobe of rat (Tanaka et al., 1997b) and frog (Alexandre et al., 2000), as well as PAC1-R mRNA in the rat pars intermedia (Shioda et al., 1997a) strongly suggests that PACAP can act as a paracrine regulator of melanotrope cell activity.
C. Effects of PACAP on the Thyroid Gland
In the human and porcine thyroid, PACAP has been shown to stimulate cAMP production and to increase thyroxine secretion (Chen et al., 1993; Kouki et al., 1997).
D. Effects of PACAP on the Gonads
The presence of PACAP and its receptors in the testis and ovary provides evidence that the peptide may operate as a local regulator of gonadal activity. In the rat testis, the concentration of PACAP is significantly reduced after hypophysectomy and is restored by FSH administration, indicating that the expression of PACAP is under the control of pituitary gonadotropins (Shuto et al., 1995). In vitro, PACAP induces a dose-dependent stimulation of testosterone secretion from isolated rat Leydig cells (Romanelli et al., 1997; Rossato et al., 1997; El-Gehani et al., 1998c) and activates or inhibits protein synthesis in spermatocytes or spermatids, respectively (West et al., 1995). In Leydig cells, PACAP activates both adenylyl cyclase and PLC through an interaction with PAC1-R (Romanelli et al., 1997). The effect of PACAP on Leydig cells may also be mediated via a novel receptor subtype coupled to a sodium channel through a pertussis toxin-sensitive G protein (Rossato et al., 1997). The effects of PACAP on protein synthesis in spermatocytes and spermatids are both mimicked by dbcAMP (West et al., 1995). In cultured Sertoli cells, PACAP increases cAMP concentration and stimulates estradiol and inhibin secretion (Heindel et al., 1992). In the epididymal epithelium, PACAP stimulates chloride secretion, which is important for sperm activation and storage (Zhou et al., 1997). The occurrence of PACAP-immunoreactive material in epididymal tubules indicates that PACAP is locally synthesized and thus may act as a paracrine regulator of sperm maturation (Zhou et al., 1997). The epithelium-derived PACAP may also stimulate epididymal spermatozoa that have lost PACAP synthesis ability (Shioda et al., 1994) but still possess PACAP binding sites (Shivers et al., 1991). In the human cavernous tissue, PACAP dose-dependently relaxes noradrenalin- and electrically-contracted preparations, suggesting that the peptide may be involved in the induction and maintenance of penile erection (Hedlund et al., 1994, 1995). In line with this finding, a stearic acid VIP conjugate has been shown to increase the copulatory activity and penile reflex in testosterone-treated, castrated rats (Gozes and Fridkin, 1992). These data suggest that PACAP and/or VIP derivates could be developed for the treatment of impotence.
In the rat ovary, most granulosa and cumulus cells from large preovulatory follicles contain both PACAP mRNA and PACAP-LI (Gräs et al., 1996). Human chorionic gonadotropin (hCG) stimulates the expression of both PACAP and progesterone receptor mRNAs (Ko et al., 1999). The peak of expression of progesterone receptor mRNA occurs 3 h after hCG treatment and the peak of PACAP mRNA only after 6 h, suggesting that progesterone receptor activation is required for PACAP gene expression (Ko et al., 1999). In support of this hypothesis, it has been shown that blockage of the progesterone receptor with the progesterone receptor antagonist ZK98299 abrogates the effect of hCG on PACAP gene expression (Ko et al., 1999). The hCG-evoked stimulation of PACAP gene transcription is abolished by cycloheximide, indicating the requirement of protein synthesis for PACAP mRNA expression (Ko et al., 1999). Exposure of cultured granulosa cells to PACAP causes a dose-dependent increase in progesterone production (Zhong and Kasson, 1994; Apa et al., 1997a,b; Gräs et al., 1999). Reciprocally, immunoneutralization of endogenous PACAP reduces progesterone formation and impairs subsequent luteinization, suggesting that PACAP plays an important role in LH-induced progesterone production during the periovulatory period (Gräs et al., 1999). Incubation of immature rat preovulatory follicles with PACAP or VIP induces a dose-dependent inhibition of follicle apoptosis (Flaws et al., 1995; Lee et al., 1999b). In luteinized granulosa cells, PACAP appears to be more potent than LH in stimulating cAMP accumulation (Richards et al., 1995;Heindel et al., 1996). In the human female genital tract, PACAP is located in nerve fibers innervating blood vessels and smooth muscle cells of the internal cervical os (Graf et al., 1995; Steenstrup et al., 1995). High concentrations of PACAP are also found throughout the human uteroplacental unit (Steenstrup et al., 1996). In vitro, PACAP induces relaxation of nonvascular smooth muscle strips from the fallopian tube and myometrium (Steenstrup et al., 1994, 1995) as well as stem villous and intramyometrial arteries (Steenstrup et al., 1996), suggesting that PACAP regulates the vascular tone in the human female reproductive tract. In placental cells, PACAP enhances cAMP formation, and hCG and IL-6 production (Desai and Burrin, 1994).
E. Effects of PACAP on the Adrenal Gland
In adrenal chromaffin cells, PACAP exerts a stimulatory action on catecholamine secretion (Watanabe et al., 1992, 1995; Isobe et al., 1993; Chowdhury et al., 1994; Guo and Wakade, 1994; Houchi et al., 1994; Perrin et al., 1995; Neri et al., 1996). PACAP also stimulates the release of brain natriuretic peptide and enkephalins, two regulatory peptides that are cosequestered with catecholamines in chromaffin granules (Babinski et al., 1996; Hahm et al., 1998), and provokes a 15-fold increase in VIP mRNA expression (Lee et al., 1999a). In vivo studies have shown that PACAP and VIP stimulate catecholamine release in anesthetized dogs through activation of dihydropyridine-sensitive L-type calcium channels (Gaspo et al., 1997;Geng et al., 1997; Lamouche et al., 1999). The effect of PACAP on catecholamine secretion was significantly enhanced during insulin-induced hypoglycemia, suggesting that the stimulatory action of PACAP on adrenochromaffin cells may contribute to normalization of glycemia (Yamaguchi and Lamouche, 1999). The effect of PACAP on catecholamine secretion is associated with activation of adenylyl cyclase and elevation of cytosolic calcium concentrations (Isobe et al., 1993, 1994; Houchi et al., 1994; Perrin et al., 1995; Chamoux et al., 1998). Incubation of adrenomedullary cells in calcium-free medium or blockage of voltage-operated calcium channels suppresses the PACAP-evoked stimulation of catecholamine secretion (Isobe et al., 1993; Houchi et al., 1995; Przywara et al., 1996; O'Farrell and Marley, 1997), indicating that the effect of PACAP on chromaffin cells is mediated through calcium influx. Concurrently, PACAP increases calcium mobilization from ryanodine/caffeine-sensitive calcium stores (Houchi et al., 1995; Tanaka et al., 1996, 1998; Shibuya et al., 1999). The effect of PACAP on catecholamine release is associated with an increase in the expression of tyrosine hydroxylase, dopamine β-hydroxylase, and phenylethanolamine N-methyltransferase (Houchi et al., 1994; Rius et al., 1994; Isobe et al., 1996; Marley et al., 1996; Tönshoff et al., 1997; Hong et al., 1998; Choi et al., 1999; Park et al., 1999). It has been shown that the stimulatory effect of PACAP on tyrosine hydroxylase activity is mediated through activation of the adenylyl cyclase/PKA transduction pathway (Marley et al., 1996). The effect of PACAP on the multiplication of adrenochromaffin cells is not yet ascertained: PACAP has been reported to stimulate proliferation of rat chromaffin cells in primary culture (Tischler et al., 1995) and to inhibit the mitogenic effect of nerve growth factor on chromaffin cells (Frödin et al., 1995; Tischler et al., 1995).
Intravenous administration of PACAP causes elevation of plasma cortisol levels in dog and calf (Edwards and Jones, 1994; Kawai et al., 1994). PACAP stimulates corticosterone and aldosterone secretion from human, rat, and chicken adrenal slices, but does not affect the release of corticosteroids from dispersed fasciculata and glomerulosa cells (Andreis et al., 1995; Neri et al., 1996; Mazzochi et al., 1997), suggesting that the response of adrenocortical cells to PACAP involves the contribution of another adrenal cell type. Exposure of human adrenal slices to the β-adrenoreceptor blockerl-alprenolol totally suppresses the steroidogenic effect of PACAP (Neri et al., 1996). Similarly, the action of PACAP on dehydroepiandrosterone and cortisol secretion by the fetal human adrenal gland is suppressed by the β-adrenoreceptor antagonist propranolol (L. Breault, L. Yon, M. Montéro, L. Chouinard, V. Contesse, C. Delarue, A. Fournier, J.G. LeHoux, H. Vaudry, and N. Gallo-Payet, submitted). Altogether, these observations indicate that, in several mammalian species, the effect of PACAP on corticosteroid secretion can be ascribed to the stimulatory action of the peptide on catecholamine secretion. In contrast, PACAP was found to stimulate corticosteroid release from dispersed bovine and frog adrenocortical cells (Yon et al., 1993b, 1994; Bodart et al., 1997). The fact that PACAP stimulates cAMP and inositol phosphate formation in bovine glomerulosa cells (Bodart et al., 1997) and calcium mobilization in individual frog adrenocortical cells (Yon et al., 1994) provides additional evidence of a direct stimulatory effect of the peptide on steroidogenesis in these two species.
F. Effects of PACAP on the Gastrointestinal Tract
Intravenous injection of PACAP to anesthetized rat stimulates secretion of saliva from the submandibular and parotid glands (Mirfendereski et al., 1997). In the ferret submandibular gland, PACAP and VIP exert a vasodilatory effect that contributes to the salivary secretory activity (Tobin et al., 1995). In the rat stomach, PACAP inhibits histamine- and pentagastrin-stimulated gastric acid secretion but has no effect on carbachol-induced secretion (Mungan et al., 1992b,1995; Li et al., 2000). In contrast, in isolated rabbit parietal cells, PACAP potentiates the response to histamine and to carbachol (Healey et al., 1998). In the gastric mucosa, PACAP has been found to stimulate histamine release from enterochromaffin cells (Lindstrom et al., 1997;Håkanson et al., 1998; Zeng and Sachs, 1998; Chen et al., 1999b; Zeng et al., 1999a) through activation of L-type calcium channels (Zeng et al., 1999b). Because histamine is a potent stimulator of chloride secretion (Helander and Keeling, 1993), this observation suggests that the effect of PACAP on gastric acid production can be accounted for, at least in part, by an indirect stimulation of enterochromaffin cells. PACAP also stimulates proliferation of gastric enterochromaffin cells through activation of the PKA, protein tyrosine kinase, and MAP kinase pathways (Lauffer et al., 1999). Intracerebroventricular injection of PACAP stimulates gastric acid secretion (Mizuta et al., 1994; Ozawa et al., 1997), suggesting that PACAP may act centrally to regulate gastric acid release possibly via an indirect mediator such as peptide tyrosine tyrosine (Guo et al., 1987). In support of this hypothesis, i.v. injection of PACAP has been shown to increase plasma peptide tyrosine tyrosine concentration (Zhang et al., 1993b). On isolated chief cells from the guinea pig stomach, PACAP increases exocytosis of zymogen granules that release pepsinogen (Felley et al., 1992). Intravenous injection of PACAP also enhances bicarbonate secretion in the duodenum (Takeuchi et al., 1997). In the distal colon, PACAP acts through cholinergic and noncholinergic neurons to evoke chloride secretions (Kuwahara et al., 1993). One of the interesting features is the superior potency of PACAP as compared with other gut neuropeptides in stimulating gastrointestinal exocrine secretions (Lauff et al., 1999).
Besides its effects on the secretory activity of exocrine and endocrine cells, PACAP induces a concentration-dependent relaxation of gastric smooth muscles (Katsoulis et al., 1996; Robberecht et al., 1998), causing a decrease of gastric motility and a delay in stomach emptying (Ozawa et al., 1999). PACAP also exerts a relaxant effect on intestinal smooth muscles in rat and in the Atlantic cod, Gadus morhua(Mungan et al., 1992a; Schworer et al., 1992; Katsoulis et al., 1993b;Grider et al., 1994; Ekblad and Sundler, 1997; Olsson and Holmgren, 2000; Pluja et al., 2000) and thus reduces the motility of the bowel (Lauff et al., 1999). In contrast, in the guinea pig small intestine, PACAP stimulates normal peristalsis and counteracts drug-induced peristaltic arrest (Heinemann and Holzer, 1999). The contractile effect of PACAP on the guinea pig ileum is mediated through presynaptic stimulation of acetylcholine and substance P release (Katsoulis et al., 1993a). In the opossum internal anal sphincter, PACAP exerts a biphasic effect, i.e., an initial contraction followed by a relaxation (Rattan and Chakder, 1997; Chakder and Rattan, 1998). The contractile but not the relaxant effect of PACAP on the anal sphincter is abrogated by a substance P antagonist, confirming that the PACAP-evoked contraction is mediated through presynaptic activation of substance P afferents (Rattan and Chakder, 1997).
G. Effects of PACAP on the Liver
It has long been known that VIP is a potent stimulator of adenylyl cyclase activity in liver cells (Waelbroeck et al., 1981). Likewise, in cultured hepatocytes, PACAP causes a dose-dependent accumulation of cAMP but does not affect inositol phosphate turnover (El Fahime et al., 1996). The fact that VIP exerts a mitogenic action on rat hepatocytes (Kar et al., 1996) strongly suggests that PACAP could be also involved in the control of liver cell proliferation. Injection of PACAP to anesthetized dogs induces a 2-fold increase of plasma glucose concentration (Sekiguchi et al., 1994). In fact, PACAP is more potent than VIP in stimulating glucose output from the perfused rat liver (Yokota et al., 1995). The hyperglycemic action of PACAP observed in vivo can be ascribed to both a direct action on hepatocytes and an indirect effect via glucagon and/or adrenaline release (Sekiguchi et al., 1994).
H. Effects of PACAP on the Pancreas
In the pancreas, PACAP-immunoreactive fibers innervate both the exocrine acini and the islets of Langerhans, as well as the small arteries of the connective tissue (Table 2; Köves et al., 1993;Tornoe et al., 1997). Electrical stimulation of the vagus nerve causes the release of PACAP from the isolated perfused pig pancreas, suggesting that PACAP may control exocrine and/or endocrine pancreatic secretions (Tornoe et al., 1997).
Intravenous injection of PACAP triggers amylase (Mungan et al., 1991;Alonso et al., 1994), pancreatic fluid, bicarbonate, and protein secretions (Naruse et al., 1992; Alonso et al., 1994; Zabielski et al., 1994; Rodriguez-Lopez et al., 1995; Onaga et al., 1996; Wheeler et al., 1997; Lee et al., 1998). PACAP also induces vasodilation and increases pancreatic blood flow, notably in the exocrine part of the gland (Carlsson et al., 1996; Ito et al., 1998). The stimulatory effect of PACAP on juice flow is inhibited by the antagonist PACAP(6–38) (Tornoe et al., 1997). Experiments conducted on isolated rat pancreatic acini have shown that PACAP exerts a direct increase on amylase and lipase secretions (Kashimura et al., 1991; Raufman et al., 1991; Schmidt et al., 1993). Coadministration of PACAP with cholecystokinin, carbachol, or bombesin to dispersed guinea pig acinar cells causes additive stimulation of amylase secretion (Kimball and Mulholland, 1996). The effect of PACAP is likely mediated via the adenylyl cyclase pathway but does not involve PLC activation or calcium mobilization (Kashimura et al., 1991; Kitagawa et al., 1995; Kimball and Mulholland, 1996). Besides its direct action on acinar cells, PACAP may also exert an indirect effect on pancreatic exocrine secretions through modulation of afferent nerve activity. In particular, PACAP has been shown to stimulate pancreatic enzyme secretion in sheep via activation of vagal cholinergic neurons (Onaga et al., 1997). PACAP also enhances electrically evoked stimulation of noradrenaline release in the canine pancreas (Yamaguchi and Fukushima, 1998), suggesting that the peptide may control juice flow through presynaptic modulation of the parasympathetic vagus nerve. Altogether, these data suggest that PACAP has to be added to the still growing list of secretagogs of the exocrine pancreas.
In the endocrine pancreas, PACAP appears to be much more potent than VIP or other regulatory peptides in stimulating pancreatic hormone secretion. In vivo administration of PACAP causes a significant increase in plasma insulin level in mice (Fridolf et al., 1992;Filipsson et al., 1998a), calf (Edwards et al., 1997), dog (Kawai et al., 1992), and humans (Filipsson et al., 1997). The stimulatory effect of PACAP on insulin release has also been documented on perfused rat and pig pancreas (Kawai et al., 1992; Yokota et al., 1993; De Stefanis et al., 1995; Bertrand et al., 1996; Tornoe et al., 1997) and on cultured islets cells (Yada et al., 1994, 1997a,b;Filipsson et al., 1998b, 1999; Davalli et al., 1999). Furthemore, pancreatic β-cells express cell-surface ectopeptidases capable of degrading PACAP (Hupe-Sodmann et al., 1997). The amplitude and kinetics of the PACAP-evoked stimulation of insulin release depends on glucose concentration in the incubation medium (Yokota et al., 1993; Bertrand et al., 1996; Edwards et al., 1997). PACAP induces a biphasic effect on insulin secretion i.e., a rapid and transient stimulation (acute phase) followed by a rebound of the secretory response (plateau phase). The phosphatidylinositol 3-kinase inhibitor wortmannin inhibits the plateau phase but not the acute phase of the PACAP-evoked insulin release (Straub and Sharp, 1996). The effect of PACAP is mediated through PAC1-R and involves activation of the adenylyl cyclase pathway (Borboni et al., 1999). Exposure of pancreatic β-cells to PACAP causes calcium influx through L-type calcium channels (Yada et al., 1997b) and the stimulatory effect of PACAP on insulin secretion is abolished by nitrendipine (Komatsu et al., 1996), indicating that activation of voltage-sensitive L-type calcium channels is involved in the insulinotropic effect of PACAP. Paradoxically, the combination of glucose, PACAP, and carbachol stimulates insulin release while being unable to elevate intracellular calcium (Komatsu et al., 1996). Incubation of isolated rat islets with specific PACAP antisera inhibits the ability of glucose to stimulate insulin release (Yada et al., 1997a; Filipsson et al., 1999), indicating that endogenous PACAP acts as a physiological regulator of pancreatic β-cell activity. PACAP is also a potent stimulator of glucagon secretion. Intravenous injection of PACAP increases plasma glucagon concentration in mice (Fridolf et al., 1992) and humans (Filipsson et al., 1997). Likewise, in the perfused rat pancreas, PACAP enhances glucagon secretion (Yokota et al., 1993). The stimulatory effect of PACAP on insulin and glucagon release is completely abolished by somatostatin (Yokota et al., 1993). In contrast, the endozepine octadecaneuropeptide (a potent inhibitor of insulin release; Tonon et al., 1997) has no effect on the PACAP-evoked insulin secretion (De Stefanis et al., 1995).
I. Effects of PACAP on the Respiratory System
The occurrence of PACAP and PACAP receptors has been reported at different levels of the airways (Tables 2 and 7). In rodents, PACAP causes relaxation of tracheal smooth muscles (Araki and Takagi, 1992; Conroy et al., 1995; Foda et al., 1995; Hiramatsu et al., 1995; Yoshihara et al., 1997) and bronchodilation (Linden et al., 1995,1997, 1999; Kinhult et al., 2000). In guinea pig, rabbit, and primate trachea precontracted with acetylcholine or potassium, micromolar concentrations of PACAP cause smooth muscle relaxation (Kanemura et al., 1993; Bhogal et al., 1994; Okazawa et al., 1998). It has also been reported that PACAP suppresses the increase in airway hyper-responsiveness induced by ozone exposure (Aizawa et al., 1999). The relaxant effect of PACAP on the trachea is mediated through activation of the cAMP/PKA (Araki and Takagi, 1992; Kanemura et al., 1993; Foda et al., 1995) and nitric oxide (NO)/cyclic guanosine monophosphate transduction pathways (Saotome et al., 1998). In addition to its potent bronchodilatory activity, PACAP is a potent stimulator of airway mucus secretions (Wagner et al., 1998; Liu et al., 1999). Owing to the bronchorelaxant properties of PACAP, synthetic analogs are currently under evaluation for their potential application in the treatment of asthma (Bolin et al., 1995; Meyer et al., 1996; Saguchi et al., 1997).
J. Effects of PACAP on the Cardiovascular System
The walls of blood vessels are richly innervated by PACAP-containing fibers (Table 2; Köves et al., 1990; Cardell et al., 1991) and a high density of PACAP binding sites is present in arteries (Table 6; Amenta et al., 1991; Nandha et al., 1991; Huang et al., 1993). PACAP, in very much the same way as VIP, is a highly potent vasorelaxant peptide (Hirata et al., 1985; Ross-Ascuitto et al., 1993;Tong et al., 1993; Ascuitto et al., 1996). Intracerebral injection of low doses of PACAP (0.1–1 nmol) produces a rapid increase in cerebral blood flow (Uddman et al., 1993; Jansen-Olesen et al., 1994; Seki et al., 1995a). Intravenous infusion of very low doses of PACAP (0.01–10 pmol/min) induces a concentration-dependent increase in blood flow and a concomitant decrease in blood pressure (Nandha et al., 1991; Ishizuka et al., 1992; Minkes et al., 1992a; Warren et al., 1992a,b; Naruse et al., 1993; Santiago and Kadowitz, 1993; Suzuki et al., 1994b;Mirfendereski et al., 1997; Whalen et al., 1999a,b,c). The most prominent effects induced by i.v. administration of PACAP are observed in the parotid and submandibular glands, the eyelids, and the nictitating membrane (Nilsson, 1994). The vasodilatory activity of PACAP also has been documented in various organs including the brain (Tong et al., 1993; Anzai et al., 1995), the eye (Nilsson, 1994; Elsas and White, 1997; Dorner et al., 1998), the pulmonary vascular bed (Minkes et al., 1992b; Cheng et al., 1993; Foda et al., 1995), the mesentery (Wilson and Warren, 1993), the pancreas (Bertrand et al., 1996; Ito et al., 1998), the testis (Lissbrant et al., 1999), the ovary (Steenstrup et al., 1994; Yao et al., 1996), the kidney (Gardiner et al., 1994), and the skin (Wallengren, 1997). In dog, administration of high doses of PACAP (3 nmol) induces a biphasic effect, i.e., a transient hypotensive response followed by a sustained hypertension (Ishizuka et al., 1992), suggesting that the action of PACAP on the vascular tone can be ascribed both to a direct vasorelaxant effect and an indirect hypertensive action mediated through the release of catecholamines. In support of this hypothesis, it has been shown that the increase in blood pressure induced by i.v. injection of PACAP in cat is abolished by the α1- and α2 -adrenoreceptor antagonist phentolamine and by adrenalectomy (Minkes et al., 1992a). The mechanism of action of PACAP on blood vessel contractility is not fully understood. The effects of PACAP on blood pressure can be ascribed, at least in part, to its relaxant activity on arterial smooth muscle cells (Huang et al., 1993; Naruse et al., 1993; Steenstrup et al., 1996; Bruch et al., 1997). PACAP increases cAMP formation in the isolated rabbit ovarian artery (Yao et al., 1996), the rat tail vein (Absood et al., 1992), and cerebral microvessels (Kobayashi et al., 1994; Wilderman and Armstead, 1997). The inhibitory effect of cAMP on smooth muscle cell contraction is well documented (Steer, 1976; Korenman and Krall, 1977; Farah, 1983). In particular, hypertension is a common manifestation in patients with cortisol excess, and glucocorticoids are known to inhibit cAMP production (Ito et al., 1994). These observations suggest that PACAP, which stimulates cAMP production in blood vessels, may have potential therapeutic value for the treatment of hypertension. PACAP modulates L-type calcium channels in vascular smooth muscle cells through the activation of both PKA and protein kinase C (PKC) (Chik et al., 1996). The action of PACAP on arteriol smooth muscle cell relaxation requires the activation of ATP- and calcium-dependent potassium channels (Bruch et al., 1997). PACAP also stimulates the release of the prostaglandin PGF2α but does not affect other cyclooxygenase metabolites (Anzai et al., 1995). The possible involvement of the endothelium in the vasodilatory activity of PACAP is still disputed: two reports indicate that the vasorelaxant effect of PACAP on the aorta and coronary arteries is endothelium-independent (Warren et al., 1991; Kastner et al., 1995) whereas another study reveals that removal of the vascular endothelium abolishes the dilatory response induced by PACAP in pulmonary arteries (Cardell et al., 1997). Some of the effects of PACAP on the vascular bed appear to be mediated through the release of vasculotropic factors. In particular, PACAP has been found to stimulate the production of vascular endothelial growth factor, which plays an important role in angiogenesis and vascular permeability (Gloddek et al., 1999).
In the heart, PACAP produces positive inotropic, chronotropic, and dromotropic effects, making it a cardiotonic candidate for treatment of heart failure. For instance, i.v. injection of PACAP in cat and sheep provokes an increase in heart rate and enhances the contractile ventricular force (Minkes et al., 1992a; Sawangjaroen et al., 1992;Sawangjaroen and Curlewis, 1994). In dog, PACAP causes transient positive followed by negative chronotropic and inotropic responses (Hirose et al., 1997b, 1998). The positive inotropic and chronotropic effects of PACAP are attributable to direct stimulation of cardiac myocytes (Suzuki et al., 1993; Runcie et al., 1995; Hirose et al., 1997a) whereas the negative chronotropic response can be ascribed to presynaptic regulation of acetycholine release from intracardiac parasympathetic nerves (Hirose et al., 1997c). In vitro studies on the isolated guinea pig heart have confirmed that the negative chronotropic effect of PACAP can be accounted for by an increase in acetylcholine release from parasympathetic neurons (Seebeck et al., 1996). In rat, the PACAP-induced tachycardia is abolished by the β-adrenoreceptor antagonist propranolol but is not affected by the ganglion blocker chlorisondamine, indicating that PACAP directly stimulates norepinephrine release from cardiac sympathetic nerve terminals (Whalen et al., 1999a).
K. Effects of PACAP on Immune Cells
On human monocytes, PACAP induces a slight increase in cAMP formation that gradually vanishes during differentiation into macrophages (Chedeville et al., 1993). On cultured mast cells, PACAP stimulates histamine secretion (Mori et al., 1994; Odum et al., 1998; Schmidt-Choudhury et al., 1999a,b) and serotonin release (Seebeck et al., 1998), suggesting that PACAP could be involved in the regulation of the inflammatory process. In mitogen-stimulated murine splenocytes, PACAP causes a dose-dependent inhibition of cell proliferation induced by concanavalin A (Tatsuno et al., 1991b). PACAP decreases chemotaxis of thymocytes and splenic lymphocytes through activation of the PKA pathway (Delgado et al., 1995; Garrido et al., 1996). In CD4+CD8+thymocytes, PACAP prevents glucocorticoid-induced apoptosis (Delgado et al., 1996b) by inhibiting Fas ligand expression (Delgado and Ganea, 2000), suggesting a possible implication in intrathymic T-cell maturation. Several studies indicate that PACAP modulates the production of cytokines by immune cells. In murine spleen cells and thymocytes, PACAP inhibits IL-10 production via both cAMP-dependent and cAMP-independent transduction pathways (Martinez et al., 1996; Wang et al., 1999). In unstimulated macrophages, PACAP and its agonist maxadilan inhibit the release of tumor necrosis factor-α (TNF-α) and increase IL-6 production through activation of PKA and PKC (Delgado et al., 1998, 1999c,g; Martinez et al., 1998a; Soares et al., 1998). In contrast, PACAP inhibits the release of both IL-6 and IL-12 as well as TNF-α from lipopolysaccharide-stimulated macrophages; this suggests that PACAP could act as a protective agent that regulates the excessive release of proinflammatory cytokines (Martinez et al., 1998a,b; Delgado et al., 1999a,c,e). Concurrently, PACAP enhances the production of the anti-inflammatory cytokine IL-10 by lipopolysaccharide-activated macrophages (Delgado et al., 1999f). Thus it appears that the anti-inflammatory activity of PACAP can be accounted for both by an inhibition of the proinflammatory cytokines IL-6 and TNF-α, and by a stimulation of the anti-inflammatory cytokine IL-10. The effect of PACAP on macrophages involves the up-regulation of B7.2 but not B7.1 gene expression (Delgado et al., 1999b,h). In addition, PACAP inhibits NO production from activated macrophages in a dose- and time-dependent manner (Delgado et al., 1999d). The release of NO is a major mechanism through which macrophages exert their cytotoxic effect against pathogens, and is also responsible for acute inflammatory diseases (Laskin and Pendino, 1995). Therefore, the inhibition of NO synthesis induced by PACAP could play a physiological role in the modulation of the inflammatory response. Inflammatory stress due to infection by various microorganisms is known to activate inflammatory regulators through the hypothalamo-pituitary-adrenocortical axis (Sternberg, 1995;Buckingham et al., 1996). Intraperitoneal administration of lipopolysaccharide stimulates PACAP-LI in the PVN, suggesting that PACAP may function as a hypothalamo-pituitary-releasing factor during acute inflammation (Hannibal et al., 1999).
L. Effects of PACAP on Bones
Immunoreactive PACAP has been detected in cartilage canals from newborn pigs (Strange-Vognsen et al., 1997), and VPAC1 receptors are expressed in human osteoblasts (Togari et al., 1997). Consistent with these observations, PACAP has been found to increase cAMP formation in mouse calvarial bones (Lerner et al., 1994) and to inhibit bone resorption by rabbit osteoblasts (Winding et al., 1997).
M. Effects of PACAP on Tumor Cells
As already noticed, PACAP and its receptors are highly expressed in a number of tumor cell lines (see section II, G and III, G). Consistent with this observation, it has been found that PACAP exerts either stimulatory or inhibitory effects on tumor cells. In the small cell lung tumor cell line NCI-H345, PACAP stimulates cell proliferation through the activation of type II binding sites (Moody et al., 1993, 1997). In rat pancreatic carcinoma AR4–2J cells, PACAP strongly increases c-fos and c-jun gene expression (Schäfer et al., 1996) and stimulates cell proliferation (Buscail et al., 1992; Douziech et al., 1998). The effect of PACAP on AR4–2J cells is mediated through activation of tyrosine kinase and phospholipase D (Morisset et al., 1995). PACAP also increases c-fos expression in lung cancer cells (Draoui et al., 1996). The fact that the PAC1-R antagonist PACAP(6–38) reduces tumor growth in nude mice transplanted with lung tumor cell (Zia et al., 1995) and breast cancer cell xenografts (Leyton et al., 1999) indicates that PACAP exerts a tonic stimulatory effect on cell proliferation. Likewise, PACAP transiently increases c-fosgene expression in prostate cancer cells in vitro, and PACAP(6–38) markedly inhibits tumor growth in mice bearing PC-3 xenografts (Leyton et al., 1998). In contrast, PACAP slackens cell proliferation of glioblastoma and colonic adenocarcinoma cells (Vertongen et al., 1996; Lelievre et al., 1998a). On neuroblastoma cell lines, PACAP exerts a biphasic effect, with stimulation of proliferation occurring at subnanomolar doses and differentiation at higher concentrations (Deutsch et al., 1993; Hoshino et al., 1993; Lilling et al., 1994;Lelievre et al., 1996, 1998b). The stimulatory action of PACAP on cell multiplication is mediated through activation of the MAP kinase cascade, independently of the PKA and PKC pathways, whereas the inhibitory action of PACAP can be accounted for by activation of PKA (Frödin et al., 1994).
In pheochromocytoma PC12 cells, PACAP stimulates tyrosine hydroxylase gene expression (Corbitt et al., 1998) and promotes neurite outgrowth (Deutsch and Sun, 1992; Lazarovici et al., 1998). In PC12 cells, PACAP, acting through type I PACAP receptors, stimulates both the PKA- and PKC-signaling cascades (Watanabe et al., 1990; Deutsch and Sun, 1992;Cavallaro et al., 1995; Kozawa et al., 1995). As a matter of fact, the action of PACAP on the differentiation of PC12 cells can be ascribed to its stimulatory effect on the PKA (Hernandez et al., 1995) and/or the PKC transduction pathways (Schadlow et al., 1992; Colbert et al., 1994). The extracellular ERK inhibitor PD98059 abrogates both PACAP-induced stimulation of ERK and neurite outgrowth (Frödin et al., 1994; Barrie et al., 1997), suggesting that activation of the MAP kinase cascade is required for initiating the differentiation of PC12 cells into sympathetic-like neurons (Traverse et al., 1992; Tanaka et al., 1997a). PACAP prevents apoptosis of PC12 cells provoked by serum depletion, through stimulation of the PKA pathway and subsequent activation of the MAP kinase cascade (Tanaka et al., 1997a). PACAP also prevents ceramide-induced apoptosis of PC12 cells by affecting signaling events downstream of the c-Jun N-terminal kinase (Hartfield et al., 1998). In addition, PACAP enhances chromogranin A gene expression (Taupenot et al., 1998), activates the transcription of the transfected NPY and proenkephalin A genes (Colbert et al., 1994;Monnier and Loeffler, 1998), and regulates genes bearing a CRE or TRE motif via an increase in cAMP and inositol phosphate formation (Schadlow et al., 1992; Monnier and Loeffler, 1998; Yukimasa et al., 1999). The chromogranin A trans-activation response induced by PACAP is subject to desensitization when the cells are pre-exposed to PACAP (Taupenot et al., 1999).
In tumor pituitary cells, PACAP modulates hormone secretion and/or cell proliferation. For instance, in the gonadotrope αT3–1 cell line, PACAP stimulates the catalytic and regulatory subunits of PKA (Garrel et al., 1997) and inhibits transforming growth factor-β-induced apoptosis in the human pituitary adenoma cell line HP75 (Oka et al., 1999). In the lactotrope 235–1 cell line, PACAP stimulates PRL release through activation of the PLC pathway and exerts mitogenic effects (Hammond et al., 1996). In the lactotrope/somatotrope cell lines GH3, nanomolar concentrations of PACAP stimulate GH and PRL release through activation of type II receptors and recruitment of voltage-gated sodium channels (Propato-Mussafiri et al., 1992; Murakami et al., 1995). In the picomolar range, PACAP increases PRL mRNA level independently of the cAMP/PKA pathway (Coleman and Bancroft, 1993; Murakami et al., 1995; Koshimura et al., 1997). In the corticotrope AtT20 cell line, PACAP mimics the effect of CRF, i.e., it stimulates adenylyl cyclase activity and triggers both POMC gene transcription and ACTH release (Koch and Lutz-Bucher, 1992a, 1995; Boutillier et al., 1994; Braas et al., 1994; Aoki et al., 1997). In the FS cell line TtT/GF, PACAP increases IL-6 secretion (Matsumoto et al., 1993). PACAP has also been found to activate human pituitary adenomas. In actively secreting adenoma, PACAP exhibits a modest stimulatory effect on ACTH, GH, or gonadotropin release (Desai et al., 1994). In nonfunctional pituitary tumors, PACAP stimulates cAMP formation and induces calcium influx through L-type calcium channels (Lania et al., 1995). Taken together, these observations suggest that PACAP/VIP receptors may be involved in the regulation of tumor cells. Development of selective PACAP agonists or antagonists should give rise to powerful pharmacological tools for the treatment of cancers (Jiang et al., 1997; Fruhwald et al., 1999).
V. Conclusion and Perspectives
Twelve years after its initial characterization, PACAP certainly appears as one of the most fascinating neuropeptides ever identified. PACAP belongs to the largest family of regulatory peptides, which encompasses several other prominent members including secretin, glucagon, GRF, and VIP. The structural and functional relationships among these paralogous peptides, as well as their receptors, provide a unique model for investigating the evolutionary processes leading to diversification of a multigene family. The primary structure of PACAP has been extremely well conserved from the sea squirt (a protochordate) to humans, indicating that this peptide must be involved in vital functions throughout the animal kingdom. As a matter of fact, PACAP has been implicated in a broad range of biological processes including reproduction, development, growth, cardiovascular, respiratory, and digestive functions, immune responses, and circadian rhythms. Whether these pharmacological responses to PACAP actually reflect physiological activities of the peptide remains a matter of speculation. To answer this fundamental question, the development of potent and selective PACAP antagonists, as well as the production of PACAP- and PACAP receptor-knockout animals, are obviously required. There is now clear evidence that PACAP exerts trophic effects on multiple types of cells but many questions remain unanswered regarding the molecular mechanisms involved in the action of PACAP on proliferation, migration, differentiation, and apoptosis. In particular, investigation of the effect of PACAP on key regulatory proteins of the cell cycle and components of the apoptotic pathways must be urgently investigated.
The beneficial effects of PACAP or PACAP antagonists in various pathological conditions such as ischemia, cancer, heart failure, and asthma will undoubtly motivate the development of new ligands, most preferably peptidomimetics, which could potentially be used as neuroprotective, antiproliferative, antihypertensive, or bronchodilatory drugs. The occurrence of multiple receptor subtypes including splice variants, which possess differential affinities for various ligands and exhibit specific tissue expression, generates hopes for the development of therapeutic agents acting on selected targets. Better characterization of the three-dimensional conformation of PACAP and analysis of the dynamic interactions of the peptide with its receptors would be instrumental for the design of such compounds. Alternatively, owing to the potential therapeutic value of PACAP receptor ligands, high-output screening of chemical libraries using cells transfected with the different PACAP receptors also should be a promising avenue for the development of novel drugs.
Acknowledgments
We thank Dr. Maı̈téMontéro, David Alexandre, and Dr. Youssef Anouar for valuable discussions and comments on the manuscript, and Sabrina Mancel for skillful secretarial assistance. This work was supported by grants from the Institut National de la Santé et de la Recherche Médicale (INSERM U413), the Ministère des Affaires Etrangères (France-Québec exchange program to A.F. and H.V.), and the Conseil Régional de Haute-Normandie. H.V. is Affiliated Professor at the Institut National de la Recherche Scientifique-Institut Armand Frappier.
Footnotes
-
↵1 Address for correspondence: Dr. Hubert Vaudry, Institut Fédératif de Recherches Multidisciplinaires sur les Peptides (IFRMP 23), Laboratoire de Neuroendocrinologie Cellulaire et Moléculaire, Institut National de la Santé et de la Recherche Médicale U413, Unité Affiliée au Centre National de la Recherche Scientifique, Université de Rouen, 76821 Mont-Saint-Aignan, France. E-mail: hubert.vaudry{at}univ-rouen.fr
Abbreviations
- ACTH
- adrenocorticotropic hormone
- CHO
- Chinese hamster ovary
- CNS
- central nervous system
- CREB
- cAMP-responsive element-binding protein
- CRF
- corticotropin-releasing factor
- EGL
- external granule cell layer
- E
- embryonic day
- ERK
- extracellular signal-regulated kinase
- FS
- folliculo-stellate
- FSH
- follicle-stimulating hormone
- GH
- growth hormone
- GnRH
- gonadotropin-releasing hormone
- GRF
- growth hormone-releasing factor
- hCG
- human chorionic gonadotropin
- IGL
- internal granule cell layer
- IL
- interleukin
- LH
- luteinizing hormone
- LI
- like immunoreactivity
- α-MSH
- α-melanocyte-stimulating hormone
- MAP kinase
- mitogen-activated protein kinase
- NO
- nitric oxide
- NPY
- neuropeptide tyrosine
- PACAP
- pituitary adenylate cyclase-activating polypeptide
- PAC1-R
- PACAP-specific receptor
- PC
- prohormone convertase
- PHI
- peptide histidine-isoleucine
- PHM
- peptide histidine-methionine
- PKA
- protein kinase A
- PKC
- protein kinase C
- PLC
- phospholipase C
- P
- postnatal day
- POMC
- proopiomelanocortin
- PRL
- prolactin
- PRP
- PACAP-related peptide
- PVN
- paraventricular nucleus
- RIA
- radioimmunoassay
- SON
- supraoptic nucleus
- TM
- transmembrane domain
- TNF-α
- tumor necrosis factor-α
- TRH
- thyrotropin-releasing hormone
- TSH
- thyroid-stimulating hormone
- VIP
- vasoactive intestinal polypeptide
- VPAC1-R
- VIP/PACAP receptor, subtype 1
- VPAC2-R
- VIP/PACAP receptor, subtype 2
- CHO
- Chinese hamster ovary
- The American Society for Pharmacology and Experimental Therapeutics
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
-
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}