


Abstract
The importance of adult neurogenesis has only recently been accepted, resulting in a completely new field of investigation within stem cell biology. The regulation and functional significance of adult neurogenesis is currently an area of highly active research. G-protein-coupled receptors (GPCRs) have emerged as potential modulators of adult neurogenesis. GPCRs represent a class of proteins with significant clinical importance, because approximately 30% of all modern therapeutic treatments target these receptors. GPCRs bind to a large class of neurotransmitters and neuromodulators such as norepinephrine, dopamine, and serotonin. Besides their typical role in cellular communication, GPCRs are expressed on adult neural stem cells and their progenitors that relay specific signals to regulate the neurogenic process. This review summarizes the field of adult neurogenesis and its methods and specifies the roles of various GPCRs and their signal transduction pathways that are involved in the regulation of adult neural stem cells and their progenitors. Current evidence supporting adult neurogenesis as a model for self-repair in neuropathologic conditions, adult neural stem cell therapeutic strategies, and potential avenues for GPCR-based therapeutics are also discussed.
I. Introduction
Only a few decades ago, scientists thought that certain cells in the body, such as cardiac myocytes and brain cells, were nonrenewable. We now know that these cells can be regenerated through specific processes involving stem cells that exist throughout life. The first evidence of adult neurogenesis was reported in the 1960s by Joseph Altman, who showed that neurons in adult rats incorporated [3H]thymidine (Altman, 1962). However, it was not until the 1990s that the idea of adult neurogenesis became widely accepted, when it was shown that the subventricular zone (SVZ1) of the lateral ventricles (Reynolds and Weiss, 1992; Richards et al., 1992) and the subgranular zone (SGZ) of the hippocampal dentate gyrus (Gage et al., 1995; Palmer et al., 1997) contain self-renewing neural stem cells (NSCs) that give rise to new neural cells. The existence of adult neurogenesis in humans was confirmed in 1998 (Eriksson et al., 1998).
G-protein-coupled receptors (GPCRs) are the largest family of membrane receptors in eukaryotes. Although the exact number of GPCRs is unknown, nearly a thousand genes encoding for GPCRs have been identified in the human genome (Takeda et al., 2002), of which approximately half are receptors for endogenous ligands. Also called heptahelical receptors, GPCRs are integral membrane proteins composed of an extracellular N terminus, seven transmembrane α-helices connected by intracellular and extracellular loops, and an intracellular C terminus. When activated, GPCRs transduce signals from outside the cell to intracellular pathways, resulting in cellular responses.
GPCRs affect the transduction of signals through heterotrimeric G-proteins, which exist bound to the inner side of the cytoplasmic membrane. G-proteins consist of three subunits, α, β, and γ, that are altered by activated GPCRs. When a ligand binds the GPCR on the cell's outside surface, it drives a conformational change, thus activating the receptor. The activated receptor then functions as a guanine-nucleotide exchange factor, exchanging GDP for GTP on the Gα subunit of the G-protein. Subsequently, the Gα-GTP subunit dissociates from the Gβγ dimer and the GPCR. Both the GTP-bound Gα and free Gβγ subunits can induce different intracellular signaling cascades and/or downstream effector proteins (e.g., adenylyl cyclases, phospholipase C, various ion channels). Because the Gα subunit possesses intrinsic enzymatic GTPase activity, it eventually hydrolyzes the GTP back to GDP, allowing Gα to reassemble with the Gβγ subunit and GPCR, returning the GPCR and G-protein to their original states. The activity of the Gα subunit is modulated by other proteins, such as the regulators of G protein signaling proteins, a type of GTPase-activating protein that accelerates GTP hydrolysis, thereby reducing the signaling (Sjögren et al., 2010). In addition, GPCRs can transduce signals without G protein involvement through G protein-independent signaling (noncanonical) pathways (Wei et al., 2003; Shenoy et al., 2006).
GPCRs are essential in the processes of neurotransmission, cell proliferation, and organ-specific function (Luttrell, 2008). Not surprisingly, GPCRs are important drug targets with at least 30% of all modern therapeutics acting at these receptors (Overington et al., 2006; Lagerström and Schiöth, 2008). The GPCR neurotransmitter systems involved in adult neurogenesis are discussed in this review. These encompass those primarily considered neuromodulators such as norepinephrine (NE), dopamine, and serotonin. Neuromodulators regulate long-range paracrine or nonsynaptic signaling through neuronal projections into the SVZ and SGZ, the two major neurogenic areas of the adult mammalian brain. Therefore, it is not surprising that the GPCRs are involved in the regulation of NSCs and their progenitors. Furthermore, increasing evidence points to the involvement of other GPCR ligands in adult neurogenesis, such as chemokines, peptide hormones, endogenous opioids, and Wnt proteins, to name a few. In this review, the general features of NSCs, methods for studying adult neurogenesis, and role of the brain vascular niche and choroid plexus in adult neurogenesis are summarized, followed by a comprehensive examination of the GPCR systems involved in modulating adult neurogenesis (Table 1). It will conclude with discussions on adult neurogenesis in pathological conditions, use of NSC therapy in the central nervous system (CNS), and the therapeutic potential of GPCR-based NSC strategies.
Role of GPCRs in adult neurogenesis
II. General Features of Neural Stem Cells and Progenitors
A. Characteristics of Neural Stem Cells in the Subventricular Zone
NSCs are defined by their ability to self-replicate and differentiate into multiple cell types found in the CNS, including neurons, astrocytes, and oligodendrocytes (Gage, 2000). Neural progenitor cells are cells that do not fully meet all of the attributes of an NSC such as neuronal pluripotency and self-renewal (Potten and Loeffler, 1990). The identity of resident adult NSCs is debated. It has been proposed that NSCs are subependymal cells (Morshead et al., 1994), astrocytes (Doetsch et al., 1999), or multiciliated ependymal cells (Johansson et al., 1999). The astrocyte theory is the most accepted and is illustrated in Fig. 1. NSCs are located in the SVZ and are represented by a subset of slowly dividing radial-like astrocytic cells called type B cells that stain positive for glial fibrillary acidic protein (GFAP). The type B can proliferate to generate rapidly dividing cells called transient amplifying progenitors (TAP or type C cells). TAP cells lose the GFAP marker associated with type B cells, but gain the Dlx2+ epitope. These TAP cells then generate migrating neuroblasts (type A cells), distinguished by the neuronal marker observed in chain-migrating neurons called polysialylated neural cell adhesion molecule (NCAM) (Bonfanti and Theodosis, 1994). This marker is important in chain migration as NCAM-deficient mice have significantly smaller olfactory bulbs (Tomasiewicz et al., 1993; Cremer et al., 1994).

Adult SVZ neurogenesis. B cells (purple) are astrocyte-like cells that serve as the SVZ stem cell (self-renews) and contact the ventricle lumen. C cells (light green) are rapidly dividing TAP cells derived through division from the B cells. C cells divide to generate committed A cells (blue), which are neuroblasts that migrate to the olfactory bulb, where they mature to become interneurons. Ependymal cells (gray) line the ventricle walls.
Molecular markers for both stem cells and their progenitors, however, are not sufficiently specific to positively identify these cell types. Common markers for NSCs and NPCs such as nestin, bromodeoxyuridine (BrdU), and doublecortin (DCX) can also be detected in other cell types found in the brain such as reactive astrocytes or cancer cells (Lendahl et al., 1990; Clarke et al., 1994; Kaneko et al., 2000; Komitova and Eriksson, 2004). Although there is no single marker for stem and/or progenitor cells, the colocalization of several markers may be used to identify (and separate) these cells. The use of several markers simultaneously with state-of-the-art flow cytometry is presently the “gold standard” in the identification and separation of cells. However, flow cytometry cannot isolate large quantities of cells for potential therapeutic uses (Preffer and Dombkowski, 2009; Tárnok et al., 2010).
The SVZ is one of only two discrete regions in the adult CNS capable of active neurogenesis. The neurons in the SVZ are continuously generated and migrate as tangentially oriented chains (Doetsch and Alvarez-Buylla, 2006) to the olfactory bulb from all rostrocaudal locations of the SVZ through a region called the rostral migratory stream (Fig. 1). The oligodendrocytes migrate laterally and dorsally into the corpus callosum, the fornix, and the fiber tracks of the striatum (Menn et al., 2006). This chain-like migration occurs in rodents and various primates, but not humans (Sanai et al., 2004). Differences between species could be due to the longer distance that separates the olfactory bulbs from the cerebrum in humans (Sanai et al., 2004) or because rodents generate more olfactory bulb neurons out of survival. Rodents produce thousands of olfactory bulb neurons every day (Alvarez-Buylla, 1997). Once these newly formed neurons reach the olfactory bulb, they migrate radially to the outer cell layers and differentiate into GABAergic interneurons (Alvarez-Buylla, 1997). In rodents, it is estimated that the time for neural progenitors in the SVZ to migrate to the olfactory bulb and differentiate is approximately 15 days (Lois and Alvarez-Buylla, 1994).
B. Characteristics of Neural Stem Cells in the Subgranular Zone
Because the hippocampus is a center of memory and learning, adult neurogenesis in the SGZ has huge potential to affect memory functions. In a distinct but parallel manner, quiescent NSCs in the SGZ are a population of radial astrocytic cells containing GFAP, Sox2, and Nestin markers (type 1 cells) (Suh et al., 2007). Sex-determining region Y-box 2 (Sox2) is a transcription factor that maintains undifferentiated state of NSCs. Nestin is an intermediate filament protein that is expressed in NSCs considered in the early stages of development. These NSCs are located within the SGZ, with their radial processes projecting through the granular cell layer. Shorter tangential processes extend along the border of the hilus and granule cell layer (Ming and Song, 2005) (Fig. 2). Similar to SVZ cells, SGZ NSCs generate actively dividing nonradial transient amplifying progenitors (type 2 cells) that lose the GFAP marker (Sox2+, Nestin+, GFAP−). Type 2 cells generate neuroblasts that are DCX-positive and differentiate and migrate locally into the glutamatergic dentate granule cell layer (Mu et al., 2010) (Fig. 2), a process that takes approximately 4 weeks (Cameron et al., 1993). New granule cells in the dentate gyrus are continuously generated locally in all mammals examined so far, including humans (Ming and Song, 2005), and survive for an extended period of at least 2 years (Eriksson et al., 1998). The adult SVZ generates the largest pool of dividing progenitor cells. In rats, 20% of the adult SVZ cell population is proliferating (Sajad et al., 2011). In the mouse SGZ, approximately 0.01% of the granule cell population is dividing (Kempermann et al., 1997; Cameron and McKay, 2001), whereas in monkeys, it is only 0.004% (Kornack and Rakic, 1999). Therefore, adult neurogenesis is considered a rare event. Adult neurogenesis has also been reported in other areas of the brain such as the neocortex and striatum, but at much lower levels (Gould et al., 1999; Gritti et al., 1999; Rietze et al., 2000; Bédard et al., 2006).

Adult SGZ neurogenesis. Astrocytes (AS; dark purple) self-renew and also divide to give rise to progenitors (D cells; blue), which differentiate into new granule cells (G cells; orange). These newly born granule cells integrate into the granule cell layer (brown G cells), where they form into mature granule cells (green). Blood vessels (BV; pink) are found close to the SGZ layer, and it is proposed that a perivascular basal lamina (BL; yellow cells) exists here similar to the ependymal cell layer found in the SVZ.
III. Adult versus Embryonic Neurogenesis
Both embryonic and adult neural stem cells are defined by their ability to self-renew, to be relatively quiescent, and to differentiate into multiple cell types. However, their cellular properties are different. Embryonic stem cells are isolated from the blastocyst stage of an embryo (Martin, 1981) and have the potential to generate the three germ layers in vitro and in vivo (Keller et al., 1993; Kennedy and Keller, 2003). Adult NSCs have a much more restricted potential and differentiate only into neurons and glia.
During development, embryonic NSCs are derived from radial glia and neuroepithelial cells lining the neural tubes. In a temporally defined sequence, subsets of neurons are generated first, followed by astrocytes and then oligodendrocytes. The sequential maturation of the different types of neurons in the developing CNS is only recently becoming uncovered (Okano and Temple, 2009). However, cortical neurogenesis is essentially completed during the embryonic period, and gliogenesis occurs largely in the first month of postnatal life (Okano and Temple, 2009). The timing of this switch from neurogenesis to gliogenesis involves both the down-regulation of neurogenic genes and the activation of proglial genes, such as the nuclear factors 1A and 1B (Deneen et al., 2006), through transcription factors (Gritti et al., 1999), epigenetic modification (Shimozaki et al., 2005), Notch signaling (Sanosaka et al., 2009), and miRNAs (Liu and Zhao, 2009).
Adult NSCs originate from the embryonic neuroepithelial radial glia, with a subset of these cells persisting in the adult in specialized regions in the brain such as the SVZ and SGZ (Doetsch, 2003). An important regulator of the temporal progression of adult neurogenesis has been found to involve miRNA-124 (Cheng et al., 2009), which first appears between the transition from TAP into neuroblasts. The number of different types of neurons generated when adult NSCs are transplanted into embryonic brains is much less compared with embryonic NSCs (Temple, 2001).
The fundamental question of why adult NSCs are more restricted in their neurogenic potential than embryonic NSCs may be due to their microenvironment, termed the stem cell “niche.” Adult SVZ cells lie in a vascular recess, a highly specialized microdomain of extracellular matrix and circulating small molecules that provide spatial and regulatory clues (Shen et al., 2008; Tavazoie et al., 2008). In addition, the ability and frequency of adult NSCs to self-renew deteriorates with age (Kuhn et al., 1996; Ahn and Joyner, 2005) and is associated with a decline of miRNA let-7b (Nishino et al., 2008).
IV. Methods for Analyzing Adult Neurogenesis
A. Bromodeoxyuridine Labeling
BrdU labeling in conjunction with the detection of various cell types and cell cycle markers is the main method of studying adult neurogenesis today (Miller and Nowakowski, 1988), replacing [3H]thymidine, which was used previously. The major advantage of BrdU is that it does not require any previous information from the cells to be labeled and can be used across species and cell types. As a thymidine analog, it is incorporated into DNA during DNA synthesis. However, there are limitations when using BrdU to study neurogenesis. One major drawback is that BrdU labels both cell division (S-phase) and DNA synthesis associated with DNA repair, gene duplication without cell division, and apoptosis (Taupin, 2007). In addition, BrdU is toxic and has been found to negatively affect the proliferation of neural progenitors in vitro (Ross et al., 2008). Using lower doses of BrdU (50 mg/kg) minimizes labeling of non–S-phase DNA synthesis and prevents BrdU toxicity (Cooper-Kuhn and Kuhn, 2002). Furthermore, detection of adult neurogenesis requires not only BrdU colocalization with neuronal markers but also proof of the absence of apoptosis. Detailed methods for analyzing neural stem and progenitor cells via BrdU are found in Kelly et al. (2009).
B. Genetic Marking with Viruses
An alternative to BrdU for assigning proliferative properties of neurogenesis is the expression of transgenes from retroviruses. The Maloney murine leukemia virus lacks nuclear import mechanisms, so viral integration and expression of DNA can occur only during mitosis. A recombinant form of the virus containing green fluorescent protein (GFP) is used. Upon viral integration, GFP is expressed, indicating cell division (Lewis and Emerman, 1994). The use of a live reporter makes retroviruses ideal for transducing a mixed culture of stem cells or targeting live cells both in vitro and in vivo, allowing direct visualization and analysis of live, recently divided cells (Stitelman et al., 2010).
Lentiviruses have also been used for the detection of dividing cells. Although retroviruses only infect cells that are actively replicating, lentiviruses will infect all cells. Lentivirus-mediated expression, besides being used to study the properties of stem cells and progenitors, has the potential to deliver therapy-based approaches in the CNS (Kouroupi et al., 2010) and to reprogram adult NSCs to non-neuronal cell lineages (Forsberg et al., 2010). Efficient suppression of transgene expression has been recently achieved using a microRNA-regulated lentiviral reporter system (Sachdeva et al., 2010). Detailed methods for either viral system are found in Peltier and Schaffer (2010).
Adenoassociated viruses have the ability to infect specific cell types based on the virus serotype capsid and act as a vector, inserting genes of interest into target cells. Adenoassociated viruses have been used to enhance or knockout expression of various regulators of neurogenesis such as brain-derived neurotrophic factor (Henry et al., 2007), cyclin-dependent kinase 5 (Lagace et al., 2008), and interleukin-4 (Kiyota et al., 2010). This DNA virus normally expresses genes transiently and is very dependent upon the injection site and serotype. Furthermore, the insert size is limited (Landgren and Curtis, 2011), and the integration frequency may be different between mitotic and postmitotic cells (Han et al., 2008).
C. Neurospheres
Neurosphere cultures have been widely used to study neural stem/progenitor cells (Gottlieb, 2002; Singec and Quinones-Hinojosa, 2008). Neurospheres refer to the sphere-forming assay used to identify stem cells based upon their functional capacity to self-renew and express pluripotency in vitro (Reynolds and Weiss, 1992). Stem cells are cultured under nonadherent conditions to form free-floating aggregates or spheres (Rietze and Reynolds, 2006). Because SGZ tissue results in more adherent cells in culture, this assay is almost exclusively used with SVZ tissue. The tissue is dissected under a microscope, then enzymatically dissociated into single cells and grown in a serum-free medium while in the presence of epidermal growth factor (EGF) (Reynolds and Weiss, 1992). A small population of cells begins to divide, initially adhering to the plate. As the sphere grows, it detaches and becomes free-floating. The majority of the cells at this point express the intermediate filament nestin, which is present in neuroepithelial cells. To demonstrate self-renewal properties, neurospheres are mechanically dissociated and recultured at the single-cell level (to give rise to clones; i.e., clonal) in the presence of EGF, a smaller subset of these cells reforming the neurospheres (referred to as secondary neurospheres). The number of single cells that reform neurospheres is a semiquantitative way to determine the number of stem cells in vivo (Pastrana et al., 2011). To demonstrate pluripotency, each neurosphere is mechanically dissociated and plated on an adherent substrate without EGF or other growth factors, initiating the differentiation process. Immunocytochemical detection of neuronal and glial markers determines whether all three cell types (neurons, astrocytes, oligodendrocytes) are present. Neurospheres can be cultured from areas outside of the SVZ, such as in the spinal cord (Vescovi et al., 1993; Weiss et al., 1996), but the addition of fibroblast growth factor (FGF) in addition to EGF is required.
Clonal neurospheres are not homogeneous but are composed of a population of stem cells, progenitors, and differentiated cells (Parker et al., 2005). When generating clonal neurospheres by plating single cells, neurospheres were observed to frequently fuse (Mori et al., 2006; Singec et al., 2006). In addition, neurosphere formation is predisposed to cells that are poised for proliferation or are actively dividing, so it is likely that it does not detect quiescent stem cells or their intrinsic properties (Pastrana et al., 2009). Cells other than true stem cells can also give rise to neurospheres (Stingl, 2009). These limitations indicate that the neurosphere assay alone cannot be used to define stem cells in vivo. Detailed methods and video on the neurosphere assay can be found at Azari et al. (2010).
D. Monolayer Cultures
Adherent monolayer culture is another method commonly used to study neural stem/progenitor cells (Gottlieb, 2002; Ray, 2008). This method is based on the same underlying principles and uses methods similar to those used in the neurosphere culture system. In both methods, the cells are cultured in defined serum-free medium with EGF and/or FGF. However, instead of plating the cells on a nonadhesive substrate to generate free-floating spheres of cells (Reynolds et al., 1992), the progenitor cells are cultured in the presence of substrates such a polyornithine, laminin, or fibronectin coated on the cultureware. Under these conditions, the cells will adhere to the coated substrates and form monolayers (Ray et al., 1993).
Monolayer cultures sidestep some of the major limitations of neurospheres, namely the poor penetration of substances into tightly packed spheres and the fact that the neurospheres often contain a heterogenous population of cells at different stages of development (Conti and Cattaneo, 2010). By allowing cells to be more isolated and continuously bathed by the medium, NSCs grown in monolayers maintain a higher degree of homogeneity. The monolayer method also has the advantage of allowing direct access to cells for pharmacological testing and electrophysiology recordings. Nonetheless, because the culture system can greatly influence the properties of the NSCs, it is important that no single in vitro assay alone be used to define stem cells in vivo. Detailed methods of the isolation and monolayer cultivation of neural precursor cells can be found at Babu et al. (2011).
V. Regulation of Adult Neurogenesis in the Brain Vascular Niche and Choroid Plexus
The vasculature is emerging as an important site in the maintenance of the stem cell niche by providing spatial cues and regulatory signals. As arterioles traverse deeper into the brain, they lose their smooth muscle layer and essentially become tubes of endothelial cells (Girouard and Iadecola, 2006). The adult SVZ and SGZ contain an extensive vascular plexus in which dividing stem cells and TAPs are located near microvessels (Palmer et al., 2000). Tight junctions between endothelial cells and perivascular astrocytes are integral components of the blood-brain barrier (Abbott et al., 2006). The perivascular end-feet of astrocytes are closely applied to the microvessel wall through the basal lamina. Small-molecule diffusion from these endothelial cells is involved in the neurogenic process (Shen et al., 2004). SVZ blood vessels promote stem cell homeostasis and self-renewal by acting as a scaffold to support the seeding of the SVZ niche (Tavazoie et al., 2008). EphA/ephrin tyrosine kinase signaling plays a crucial role in the maintenance of the brain vascular system. In a recent study, Ephrin-A5-deficient mice display narrower hippocampal capillaries (Hara et al., 2010). These mice also have impaired SGZ-mediated neurogenesis.
Endothelial cells in the SVZ vascular niche may allow key small molecules such as vascular endothelial growth factor and FGF2 to diffuse to the NSCs and TAPs, which are established signals in the neurogenic process (Leventhal et al., 1999; Shen et al., 2004). In addition to small molecules, hormones and their antagonists that cross the blood-brain barrier may also have direct access to the SVZ. This provides a potential mechanism for GPCR-based therapeutics to regulate neurogenesis.
The choroid plexus is composed of modified ependymal and epithelial cells located in each of the four ventricles that produce cerebrospinal fluid (CSF). The choroid plexus is also composed of many capillaries and set apart from the ventricles by a layer of epithelial cells. Liquid derived from the plasma filters through these cells and becomes the CSF. The choroid plexus also removes waste products, foreign substances, and excess neurotransmitters from the CSF (Wright and Saito, 1986).
GPCRs expressed in the choroid plexus may have direct or indirect actions on the SVZ, depending upon the innervation and local release of the hormone. The sympathetic nerves innervate the choroid plexus almost exclusively by the superior cervical ganglion (Edvinsson et al., 1974; Lindvall et al., 1978b). Cholinergic and peptidergic systems also innervate the choroid plexus (Lindvall et al., 1978a; Ando et al., 1986). Serotoninergic fibers are not found in the choroid plexus (Napoleone et al., 1982) but fibers from the dorsal raphe nucleus end at the ependymal surface and release serotonin in the CSF.
GPCRs found in the choroid plexus include the adhesion receptor GPR124, which is up-regulated after brain injury (Pickering et al., 2008). Other GPCRs confirmed or suspected to be expressed in the choroid plexus include the serotonin 5-HT2C (Pazos and Palacios, 1985; Conn et al., 1986; Labasque et al., 2008), 5-HT6 (Roberts et al., 2002), cannabinoid (Ashton et al., 2004; Suárez et al., 2010), histamine H4 (Maślińska et al., 2009), orexin A (Tafuriet al., 2009), sphingosine-1-phosphate1–5 or EDG1–5 (Caballero et al., 2009), P2Y (Johansson et al., 2007), secretin (Siu et al., 2006), melanocortin type 2 (Nimura et al., 2006), corticotropin-releasing factor 2α (Chen et al., 2005), metabotropic glutamate (Gillard et al., 2003), somatostatin (Katz et al., 2002), vasopressin 1b (Zemo and McCabe, 2001), adrenomedullin (Juaneda et al., 2001; Kobayashi et al., 2001), luteinizing hormone/chorionic gonadotropin (Lei and Rao, 2001), kinin B1 (Mahabeer et al., 2000), dopamine (Mignini et al., 2000), CXCR3 (Van Der Meer et al., 2001), endothelin B (Nakagomi et al., 2000), and the α1-adrenergic (Papay et al., 2006; Gupta et al., 2009).
VI. G-Protein-Coupled Receptor Regulation of Adult Neurogenesis
A. Adrenergics
Adrenergic receptors (ARs) mediate the sympathetic nervous system by binding the endogenous catecholamines epinephrine and NE. Raymond Ahlquist (Ahlquist, 1948) introduced the initial concept of different AR subtypes (α and β). Nine AR subtypes (α1A, α1B, α1D, α2A, α2B, α2C, β1, β2, and β3) are now characterized. These receptors are all activated by the same catecholamines but result in various downstream effects (Bylund, 2005).
The adult hippocampus receives strong noradrenergic innervation from the locus ceruleus (Loy et al., 1980), and it is known that the early maturation of neural progenitor cells involves NE (Kärkkäinen et al., 2009). Pharmacological agents that increase NE levels can enhance hippocampal neurogenesis (Malberg et al., 2000) and the survival and differentiation of granule cells (Rizk et al., 2006) and olfactory bulb neurons (Bauer et al., 2003; Veyrac et al., 2005). NE was shown to activate self-renewal and produce multipotent neural precursors in the adult mouse hippocampus (Jhaveri et al., 2010). Intrahippocampal injection of a β3-AR agonist or systemic injection of isoproterenol, a β-AR nonselective agonist, increased proliferation and the number of nestin/GFAP double-positive neural precursors (Jhaveri et al., 2010). Studies using a noradrenergic neurotoxin, N-(2-chloroethyl)-N-ethyl-2-bromo benzylamine hydrochloride (DSP-4), suggest that NE regulates proliferation but not survival or differentiation of precursors in the adult rat hippocampus (Kulkarni et al., 2002). However, this neurotoxin has been shown to increase norepinephrine release (Kask et al., 1997) and regenerative sprouting (Fritschy and Grzanna, 1992) as compensatory mechanisms.
Previous studies indicate that α1-ARs increase proliferation of embryonic neuroepithelial cells, suggesting that this subtype might influence adult progenitor proliferation (Popovik and Haynes, 2000; Kulkarni et al., 2002). α1-ARs stimulate proliferation (Hiramoto et al., 2006) and migration (Hiramoto et al., 2008) and protect against stress-induced death of mouse embryonic brain-derived neural progenitor cells through a caspase 3/7-independent mechanism in vitro (Ohashi et al., 2007). We have shown that α1A-AR subtype stimulation increases neurogenesis and gliogenesis in adult mice in vivo (Gupta et al., 2009). Adult mice expressing wild-type (WT) α1A-ARs tagged with enhanced GFP, mice with constitutively active mutant receptors under the control of a large fragment of the endogenous promoter, or normal WT mice treated with the α1A-AR selective agonist cirazoline, display increased BrdU incorporation in both the SVZ and SGZ, increased migration of α1A-AR positive cells, and increased numbers of Brdu/Nestin double-positive cells. In neurospheres derived from these mice, α1A-AR stimulation increases neuron and oligodendrocyte differentiation through a phosphoinositide 3-kinase (PI3K) survival mechanism that selectively increased the apoptosis of astrocytes.
α2-ARs exist both on target neurons receiving noradrenergic innervation and as α2-autoreceptors that decrease NE release from the presynaptic terminal. In the adult, the α2-AR agonists clonidine and guanabenz decrease proliferation of hippocampal progenitors with no effect on survival or differentiation (Yanpallewar et al., 2010). Adult hippocampal progenitors in vitro express all the α2-AR subtypes that decrease neurosphere frequency and BrdU incorporation, effects that are blocked with the α2-AR antagonist yohimbine. This study also observed no changes in hippocampal progenitor proliferation in any of the α2-AR knockout (KO) mice or with α2-AR antagonists, suggesting that α2-ARs induce a stimulatory, but not a basally, tonic inhibitory effect on adult SGZ progenitors. In contrast, another study (Rizk et al., 2006) reported enhanced progenitor survival after long-duration treatment with the α2-AR antagonist dexefaroxan.
B. Cannabinoids
The cannabinoid system plays a major role in the CNS and is emerging as a key regulator of adult neurogenesis, neuronal cell fate, and neuroprotection (Gowran et al., 2011). The two currently known cannabinoid (CB) receptors, CB1 and CB2, are both GPCRs. The CB1 receptors are expressed primarily in the CNS, whereas CB2 receptors are found mainly in hematopoietic and immune cells. Endocannabinoids are the endogenous ligands of CB receptors. Endocannabinoids promote long-lasting adult plasticity in a few areas of the brain, particularly in the dentate gyrus. In contrast, most drugs of abuse decrease adult hippocampal neurogenesis (Eisch et al., 2000; Abrous et al., 2002; Nixon and Crews, 2002),
In murine NPCs, anandamide, a nonselective endogenous cannabinoid, affects fate determination in vitro by promoting first glial and then neuronal differentiation without effects on survival or apoptosis (Soltys et al., 2010). These effects were shown to depend upon the phosphorylation of a cAMP-responsive element binding protein. Activation of diacylglycerol lipases involved in the synthesis of 2-arachidonoylglycerol, as well as CB1/CB2 receptor agonism, increases proliferation of cultured NSCs or hippocampal progenitors, whereas CB receptor antagonism and CB receptor-deficient cells result in inhibited proliferation (Aguado et al., 2005; Jiang et al., 2005; Palazuelos et al., 2006; Goncalves et al., 2008). However, the CB2 receptor-mediated effect is more pronounced in the SVZ of older mice, suggesting that CB2 receptor agonists may counteract the natural decline in adult neurogenesis that is associated with aging (Goncalves et al., 2008; Marchalant et al., 2008, 2009a,b).
Neurosphere proliferation is regulated by both CB1 and CB2 receptor agonism through PI3K/AKT signaling (Molina-Holgado et al., 2007). Endocannabinoids are also thought to be involved with neuronal survival functions through PI3K/AKT phosphorylation (Galve-Roperh et al., 2008; Maison et al., 2009) and through inhibition of glutamate neurotoxicity (Galve-Roperh et al., 2008). In addition, cannabinoids oppose the antineurogenic effects of neuronal nitric oxide (Kim et al., 2006b).
The plant-derived extracts Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD) produce differential effects on spatial learning and neurogenesis when fed to nestin-GFP reporter mice (Wolf et al., 2010). CBD, working through the CB1 receptor, does not impair learning but increases adult hippocampal neurogenesis, whereas THC reduces learning without affecting neurogenesis. CB1 receptor-deficient mice have reduced adult neurogenesis in both the SVZ and SGZ (Jin et al., 2004b). Because of the lack of psychoactive effects, CBD may represent a more promising candidate for therapeutic applications than THC (Pertwee, 2009). There is one report that the endogenous endocannabinoid anandamide and its analog methanandamide may inhibit the differentiation of cortical neuron progenitors to mature neurons and reduce adult hippocampal neurogenesis through attenuation of the Rap1/B-Raf/ERK pathway (Rueda et al., 2002).
It is possible that exo- and endocannabinoids may have differential effects on hippocampal neurogenesis as a result of their full versus partial agonism at CB receptors (Fride and Mechoulam, 2003). The CB1 receptor and the endocannabinoid-inactivating enzyme fatty acid amide hydrolase are expressed, in vivo and in vitro, in postnatal radial glia, adult nestin/GFAP-positive progenitor SVZ cells (Aguado et al., 2006; Arévalo-Martín et al., 2007), and nestin/sox2-positive cells (Aguado et al., 2005; Mulder et al., 2008). In both cell culture and in postnatal CB1 receptor-deficient mice, CB1 receptors seem to regulate both progenitor proliferation and astrogliogenesis (Aguado et al., 2006). CB1 receptor activation increases the number of Oligo2-positive cells, whereas CB2 receptor activation increases polysialylated NCAM expression, both found on oligodendrocytes (Arévalo-Martín et al., 2007), through survival mechanisms involving PI3K/AKT (Molina-Holgado et al., 2002). Therefore, diseases and disorders characterized by glial deficiency potentially could be modulated through treatment with cannabinoids.
C. Chemokines (Stromal Cell-Derived Factor 1/CXC Chemokine Receptor Type 4)
Chemokines are small cytokines or proteins that are categorized into four groups. CXC chemokines (or α-chemokines) promote the migration of neutrophils and lymphocytes. CC chemokines (or β-chemokines) induce the migration of monocytes, natural killer cells, and dendritic cells. C chemokines (or γ-chemokines) attract T-cell precursors to the thymus, and CX3C chemokines (or δ-chemokines) serve as chemoattractants and adhesion molecules (Bonecchi et al., 2009).
CXCL12 [also known as stromal cell-derived factor 1 (SDF-1)] is the only known physiological ligand for the chemokine receptor CXCR4. It is a small chemokine protein of 8 to 13 kDa but is a well known mediator of neural progenitor cell migration during development. Mice that lack either the CXCR4 receptor or SDF-1 show abnormal development of the granule layer of the cerebellum (Ma et al., 1998; Zou et al., 1998) and the dentate gyrus of the hippocampus (Bagri et al., 2002; Lu et al., 2002), which suggests that this receptor system is a critical regulator of neurogenesis. During development, SDF-1 is expressed by cells lining migratory paths and by cells located at the end of migratory paths. The migrating cells themselves express the receptor CXCR4 (Ma et al., 1998; Zou et al., 1998). Anatomical studies using in situ hybridization and immunohistochemistry to localize chemokine receptors in the brain have demonstrated that they are expressed in the adult SGZ, SVZ, and olfactory bulb (Tran and Miller, 2005). Cells expressing these receptors have also been shown to express nestin and the TLX (tailless) gene—both markers for neural progenitors (Shi et al., 2004). In adult neural progenitor cells, SDF-1 induces migration and differentiation by increasing the expression of matrix metalloproteinases 3 and 9 (Barkho et al., 2008). In the postnatal brain, SDF-1 and CXCR4 are expressed in granule neurons and dividing neural progenitors in the dentate gyrus (Lu et al., 2002; Banisadr et al., 2003; Bhattacharyya et al., 2008). SDF-1 plays a novel role as a neurotransmitter in the dentate gyrus and increases the strength of GABAergic inputs to the pool of dividing neural progenitors (Bhattacharyya et al., 2008).
Excitation-neurogenesis coupling is the process by which neuronal electrical activity affects the production of new neurons (Deisseroth et al., 2004). For example, GABAergic inputs and activation of GABAA receptors has been shown to promote neuronal differentiation and development in the adult hippocampus (Tozuka et al., 2005; Ge et al., 2006). Excitation-neurogenesis is the tonic activation of CXCR4 by SDF-1 in newly formed granule cells. It is suggested to be essential for neurogenesis-dependent long-term memory in the adult hippocampus (Kolodziej et al., 2008).
Human neural progenitor cell proliferation is regulated through the CXCR4/AKT-1/Forkhead box O3a signaling pathway (Wu et al., 2009) and changes in cell cycle proteins that affect neuronal survival (Khan et al., 2003). AKT-1 promotes proliferation by interacting with 14-3-3 proteins that sequester p21 or by increasing the expression of cyclin D proteins (Muise-Helmericks et al., 1998; Zhou et al., 2001). In addition, AKT-1 phosphorylates and inhibits the winged-helix family of transcription factors (Forkhead box O3a), which are negative regulators of cell cycle progression (Brunet et al., 1999; Nakamura et al., 2000; Brunet et al., 2001). There are contrasting reports as to whether SDF-1 promotes quiescence (Krathwohl and Kaiser, 2004) or proliferation (Gong et al., 2006). One study that used primary cells from rat cortex found that SDF-1 promotes neural progenitor cell proliferation via the ERK1/2 and PI-3 kinase pathways (Gong et al., 2006), whereas the other study observed that SDF-1 inhibits growth and differentiation and induces quiescence of human NPCs in isolated intermediate progenitor cells, neurospheres, and hippocampal slice cultures (Krathwohl and Kaiser, 2004). Species differences or preparation variance are the most likely cause of contrasting results.
D. Dopamine
Dopamine is a catecholamine that is synthesized by neurons most commonly found in the substantia nigra, ventral tegmental area, and hypothalamus. Dopamine neurological function encompasses roles in punishment and reward, mood, sleep, attention, and learning (Lima et al., 2009). Dopamine receptors are classified into two groups, D1-like (D1 and D5) and D2-like (D2, D3, and D4), on the basis of their structure and interactions with different G-proteins. D1-like receptors activate adenylyl cyclase through Gs, whereas D2-like receptors inhibit adenylyl cyclase activity through Gi (Holmes et al., 2004).
Dopamine receptors are known to be expressed in neurogenic regions. Dopamine receptors, particularly D3, are highly expressed in the developing brain in areas especially active in neurogenesis and persist in the adult SVZ (Diaz et al., 1997). In the adult rodent brain, both D1- and D2-like receptors have been identified in SVZ-derived neurospheres (Coronas et al., 2004), specifically in β-III immunopositive neuronal cells. mRNA expression and in vivo studies have demonstrated a particularly strong D3 receptor presence in embryonic and adult SVZ neurogenic regions (Diaz et al., 1997; Van Kampen et al., 2004). Fluorescence-activated cell sorting of adult mouse SVZ cells using hGFAP-GFP and DCX-GFP mice showed that TAP cells express D3 receptors, whereas stem cell-like astrocytes and neuroblasts do not (Kim et al., 2010).
Dopamine is known to enhance adult neurogenesis (Winner et al., 2009). Depletion of dopamine in rodents decreases precursor cell proliferation in both the subependymal zone and the SGZ. Proliferation in the SVZ diminishes significantly after dopaminergic deafferentation. However, the number of neural progenitor cells expressing the proneuronal cell fate determinant Pax-6 increases in the SVZ. Survival and quantitative cell fate analysis of newly generated cells revealed that 6-hydroxydopamine (6-OHDA) lesions induce opposing effects in the two different neurogenic regions of the brain: a transient decrease in the granule cell layer causes a sustained increase of newly generated neurons in the glomerular layer (Winner et al., 2006). Destruction of dopamine neurons in the substantia nigra and ventral tegmental areas reduces the number of proliferating neural precursors in the SVZ by 40% (Baker et al., 2004). Inhibition of dopaminergic transmission in adult rats in vivo using the D2-like antagonist haloperidol led to inconsistent results (Wakade et al., 2002; Kippin et al., 2005). However, systemic treatment of either normal or dopamine-depleted rats with the D2/3-like agonists ropinirole or 7-hydroxy-N,N-di-n-propyl-2-aminotetralin significantly increases precursor cell proliferation in the SVZ (Höglinger et al., 2004; Van Kampen et al., 2004; Van Kampen and Robertson, 2005), resulting in increased differentiation of neurons. These studies identify dopamine D2-like receptors as a regulator of adult neurogenesis with implications for the potential use of endogenous neural precursors in cell replacement strategies for Parkinson's disease (PD).
There are conflicting reports on the ability of dopamine to promote neurogenesis, although these studies are in the minority. One study that explored 7-hydroxy-N,N-di-n-propyl-2-aminotetralin on human and murine NPCs derived from the fetal midbrain found no effects on proliferation, survival, or neurogenesis (Milosevic et al., 2007). Another study showed that repetitive cocaine administration decreases proliferation in the adult rat hippocampus with no effect on differentiation or survival (Yamaguchi et al., 2004; Domínguez-Escribà et al., 2006). It is possible that these discrepancies in the literature are due to activation of distinct subtypes of dopamine receptors, particularly in the SGZ, at different developmental stages. A recent study revealed that dopamine is particularly effective in modulating the function of adult newborn neurons in the SGZ but not in mature dentate granule cells (Mu et al., 2011). It is also possible that optimum dopaminergic modulation of neurogenesis requires simultaneous activation of D1- and D2-like receptors, shown to rescue proliferation in the 6-OHDA animal model with significantly greater increases in neuronal differentiation in the olfactory bulb than with levodopa alone (O'Keeffe et al., 2009a).
D2-like receptors are thought to promote adult neurogenesis. In vivo, D3 receptor antagonism reduces the numbers of newborn neurons that reach the olfactory bulb and decreases progenitor cell proliferation but does not change the number of BrdU-retaining (stem) cells. No effect of the D1-like agonist 2,3,4,5-tetrahydro-7,8-dihydroxy-1-phenyl-1H-3-benzazepine (SKF 38393) on cell proliferation is observed (Höglinger et al., 2004). These studies suggest that D2-like receptors (i.e., D3 receptors) are expressed on transit-amplifying progenitor cells but not SVZ stem cell-like astrocytes (Kim et al., 2010).
A variety of factors are known to influence dopamine-induced neurogenesis. It has been proposed that the mechanism of D2 receptor-mediated adult neurogenesis in mice involves ciliary neurotrophic factor (CNTF) (Yang et al., 2008b). Dopaminergic denervation in adult mice reduces CNTF mRNA by approximately 60%, whereas systemic treatment with the D2 receptor agonist quinpirole increases CNTF mRNA in the SVZ and SGZ. Quinpirole acted on postsynaptic receptors in that it reversed the reduced proliferation seen after dopaminergic denervation in wild-type mice. In vitro studies using human NPCs show that treatment with NMDA during proliferation and differentiation increases the amount of tyrosine-hydroxylase-immunopositive cells, which is reversed by memantine (Wegner et al., 2009). These results suggest that N-methyl-d-aspartate glutamate receptors in differentiating human NPCs are important regulators of dopaminergic neurogenesis in vitro. Another factor involved in regulating dopamine-induced proliferation in the SVZ is EGF, but not FGF, in a PKC-dependent manner (O'Keeffe et al., 2009b). In vivo dopamine depletion decreases proliferation in the SVZ concomitant with reduced EGF and is reversed by administration of levodopa. In addition, EGFR-positive cells are depleted in patients with Parkinson's disease (O'Keeffe et al., 2009b).
E. Glutamate
Glutamate is the major excitatory neurotransmitter of the CNS. The glutamate system mediates its effects through both ionotropic ligand-gated ion channel receptors and metabotropic glutamate receptors (mGluR), which couple to G proteins. The main glutamatergic input to the hippocampal dentate gyrus is from the entorhinal cortex via the perforant path (Collingridge and Lester, 1989). Most of the actions of glutamate on adult neurogenesis have been ascribed to its ionotropic receptors. The effects of the mGluRs on adult neurogenesis are unclear and conflicting. There are eight known subtypes of mGluRs divided into three groups: group 1 (mGluR1 and -5), group 2 (mGluR2 and -3), and group 3 (mGluRs 4, 6, 7, and 8).
The subtypes mGluR3, mGluR4, and mGluR5 are expressed in postnatal NSCs (Canudas et al., 2004; Di Giorgi Gerevini et al., 2004, 2005). Subtype-specific effects of mGluRs on adult neurogenesis remain unclear. mGluR1 antagonism, but not mGluR4/5 blockade, reduces cell proliferation in hippocampal organotypic slices, which suggests that mGluR1 receptors may facilitate neurogenesis (Baskys et al., 2005). In cultured mouse NPCs, mGlu2/3R or mGlu5R blockade reduces cell proliferation and survival, whereas mGluR5 activation increases neurogenesis (Di Giorgi-Gerevini et al., 2005). Additional studies have shown that treatment with the mGluR2/3 blocker (1R,2R,3R,5R,6R)-2-amino-3-(3,4-dichlorobenzyloxy)-6-fluorobicyclo[3.1.0]hexane-2,6-dicarboxylic acid (MGS0039) enhances cell proliferation in vivo (Yoshimizu and Chaki, 2004). It was recently shown that mGluR3 activation inhibits astrocytic differentiation of SVZ-derived NSCs through a mechanism involving bone morphogenetic protein receptor signaling (Ciceroni et al., 2010) and that treatment with the novel mGluR2 agonist (2R,4R)-4-aminopyrrolidine-2,4-dicarboxylate inhibits diffuse brain injury-induced neurogenesis (Feng et al., 2011). There is evidence suggesting a link between mGlu4R function in cerebellar granule cell neuroprogenitors and medulloblastomas and mGlu3R-mediated proliferation and survival of forebrain neural stem/progenitor cells and malignant gliomas [clearly, more study is needed to determine the significance of glutamate in regulating adult neurogenesis (for review, see Melchiorri et al., 2007)].
F. Lysophosphatidic Acid and Sphingosine 1-Phosphate
Lysophosphatidic acid (LPA) and sphingosine-1-phosphate (S1P) are extracellular bioactive phospholipids that have recently emerged as important influences on normal nervous system development. These ligands act through at least five specific GPCRs for each system, LPA1–5 (Chun et al., 2002; Meyer zu Heringdorf and Jakobs, 2007) and S1P1–5 (Rosen et al., 2009).
During embryonic development, LPA regulates cortical growth, proliferation, differentiation, and cell survival (Kingsbury et al., 2003). In embryonic tissue studies, LPA induces proliferation of murine cortical neuroblasts in vitro (Contos et al., 2000) but not in vivo (Kingsbury et al., 2003). However, it has no proliferative effect on hippocampal neural progenitor cells of the embryonic rat (Harada et al., 2004). These contrasting results could suggest that the effects of LPA are dependent upon the tissue source, species, and developmental stage of the subject (for review, see Pébay et al., 2007).
During development, LPA1 is expressed in neural progenitor cells, suggesting a regulatory function in neurogenesis (Hecht et al., 1996). In the embryonic mouse brain, LPA1 is expressed in bands of cells adjacent to several ventricles and near the lateral ventricle in the ventricular zone of the neocortex, locations that are coincident with neurogenesis (McGiffert et al., 2002).
In human embryonic stem cell-derived neurospheres, all five of the LPA receptors are expressed. LPA specifically inhibits the differentiation of NSCs toward neurons without affecting proliferation, whereas it maintains the differentiation of NSCs toward astrocytes (Dottori et al., 2008). These effects are not blocked by pertussis toxin, which inhibits Gi signaling, and are only partially inhibited by (S)-phosphoric acid mono-{2-octadec-9-enoylamino-3-[4-(pyridin-2-ylmethoxy)-phenyl]-propyl} ester (VPC32183), an antagonist of the LPA1 and LPA3 receptors. In contrast, these effects are fully blocked by a combination of PI3K/AKT and Rho/Rock inhibitors, suggesting the involvement of LPA receptors other than LPA1/3.
LPA is also known to increase adult neurogenesis by promoting the differentiation of cells toward neuronal lineages (Svetlov et al., 2004). In neurospheres obtained from mouse postnatal forebrain, LPA induces proliferation of cells coexpressing Sca-1 and AC133, markers of primitive hematopoietic and NSCs, along with the LPA receptor subtypes 1 to 3 (Svetlov et al., 2004). These effects can be blocked by diacylglycerol-pyrophosphate, an antagonist of LPA1 and LPA3 receptors. In adult mice, LPA1 is thought to promote adult neurogenesis. Although 50% of LPA1-null mice die perinatally, a variant that arises spontaneously after colony expansion, termed maLPA1, exhibit reduced ventricular zone, altered neuronal markers, and increased cortical death (Estivill-Torrús et al., 2008). Subsequent examination of these mice shows defects in proliferation, differentiation, and survival of newly formed neurons in the SGZ (Matas-Rico et al., 2008). In addition, conditions known to induce neurogenesis, including enriched environment and voluntary exercise, are impaired in the maLPA1 mice. Analysis of trophic factors in maLPA1-null mice demonstrate alterations in brain-derived neurotrophic factor and insulin growth factor 1 levels after enrichment and exercise (Matas-Rico et al., 2008). These mice also display increased anxiety and spatial memory defects (Santin et al., 2009).
S1P1 is expressed in bands of cells adjacent to several ventricles and near the lateral ventricle in the embryonic mouse brain (McGiffert et al., 2002). The expression pattern of S1P1 also encompasses the hippocampal primordia, olfactory bulb, and ganglionic eminence (McGiffert et al., 2002). In humans, embryonic SC-derived NSCs express S1P1,3, which induces proliferation and morphological changes of neural progenitor cells (Harada et al., 2004; Dottori et al., 2008). LPA1 and S1P5>1–3 are expressed in embryonic and adult rodent OPCs (Dawson et al., 2003; Terai et al., 2003; Jaillard et al., 2005; Novgorodov et al., 2007), whereas fetal human OPCs express higher levels of S1P1 (Miron et al., 2008). OPC maturation requires first the formation of processes involved in OPC migration, the actual migration of OPCs and is complete upon its myelination of neurons (Baumann and Pham-Dinh, 2001). Both LPA and S1P act on OPCs and inhibit oligodendrocyte maturation (Dawson et al., 2003; Jaillard et al., 2005). S1P5 is preferentially expressed in OPCs and is responsible for the inhibition of their migration in rodents (Novgorodov et al., 2007). In human OPCs, S1P5,3 activation with the sphingosine analog fongolimod (FTY720), a systemic immunomodulatory therapy for multiple sclerosis, has a biphasic effect on maturation. Short-term (1 day) FTY720 treatment causes initial process retraction, whereas a 2-day treatment stimulates process formation and increases cell survival in a S1P1-ERK-dependent manner (Coelho et al., 2007; Miron et al., 2008). There are also reports that S1P signaling cross-talk with neurotrophin-3 stimulates proliferation and survival of OPCs through a cAMP-response element-binding protein (Saini et al., 2004, 2005).
G. Melatonin
Melatonin is a widespread, naturally occurring compound. Many of its physiological effects are mediated through its activation of the GPCRs, MT1 and MT2. Its secretion into the blood from the pineal gland varies in a daily cycle, thereby allowing the regulation of sleep and circadian rhythm (Dubocovich et al., 2003). Adult hippocampal neurogenesis is affected by circadian rhythms and sleep deprivation (Holmes et al., 2004; Guzman-Marin et al., 2005), thus implying a significant role for melatonin. In vitro, melatonin increased the number of new neurons differentiated from adult mouse hippocampal neural precursor cells by promoting cell survival without affecting proliferation (Ramírez-Rodríguez et al., 2009; Rennie et al., 2009). Although a previous study revealed increased proliferation in the dentate gyrus of maternally separated rats seems contradictory, this study only measured BrdU incorporation without the use of neurogenetic cell markers (Kim et al., 2004). Melatonin's effect on neuronal cell survival were recapitulated in vivo by application of exogenous melatonin (8 mg/kg) and demonstrated antidepressant behavior in mice (Ramírez-Rodríguez et al., 2009). Melatonin also increases neuritogenesis and dendritogenesis with agonist treatment, leading to greater complexity of the dendritic tree (Ramirez-Rodriguez et al., 2011). Melatonin has also been shown to ameliorate the irradiation-induced decline in adult hippocampal neurogenesis, suggested to occur through its ability to scavenge free radicals (Manda et al., 2009). This benefit may be useful to combat the side effects of brain radiotherapy.
H. Muscarinic
While cholinergic activity plays a significant role in adult neurogenesis, most of these effects are documented through nicotine and the acetylcholine receptor ion channels (for review, see Campbell et al., 2011). Little is known about the role of cholinergic receptors in the GPCR system and their role in adult neurogenesis. Through immunohistochemistry and reverse transcription-polymerase chain reaction, M1 and M4 receptors were expressed in immature neurons in the adult mouse olfactory bulb or dentate gyrus (Kaneko et al., 2006) or those that colabeled with BrdU in the rat subgranular zone (Mohapel et al., 2005). The M2, M3, and M4 muscarinic receptors are expressed in embryonic rat neuroepithelial cells, and muscarinic agonists increased proliferation and differentiation of neurons (Ma et al., 2000). The nonselective muscarinic agonist, oxotremorine could alleviate the stress-induced decrease in the proliferation, survival, and differentiation of progenitor cells in the hippocampus (Veena et al., 2011). However, in contrast to an earlier study, muscarinic blockage with scopolamine only affected survival of BrdU+ cells without changing neural proliferation or differentiation in the unstressed adult rat hippocampus (Kotani et al., 2006). These results could indicate that muscarinic activity becomes important in adult hippocampal neurogenesis only under stress conditions.
I. Opioids
The classic opioid receptor system modulates many different aspects of physiology but is most known for its control of nociception (Fields, 2004) and reward-related behavior (Nestler, 2004). Four major subtypes of opioid receptors have been cloned: μ-opioid receptors (MORs), δ-opioid receptors (DORs), κ-opioid receptors (KORs), and nociception receptors (Waldhoer et al., 2004). All are GPCRs and are activated by endogenous opioid peptides.
Endogenous opioids and exogenously administered synthetic opiate agonists acting on the MORs, DORs, and KORs have been shown to affect proliferation and differentiation in various embryonic neural cell types. In embryonic NSCs, stimulation of DORs using (+)-4-[(αR)-α-(2S,5R)-4-allyl-2,5-dimethyl-1-piperazinyl)-3-methoxybenzyl]-N,N-diethylbenzamide (SNC80) promotes neuronal differentiation through PI3K/PKC/Ca2+/calmodulin-dependent protein kinase II/mitogen-activated protein kinase kinase, but MOR agonism ([d-Ala2,N-Me-Phe4,Gly5-ol]enkephalin) or KOR stimulation [trans-(±)-3,4-dichloro-N-methyl-N-[2-(1-pyrrolidinyl)cyclohexyl]benzeneacetamide (U50,488H)] have no effect (Narita et al., 2006). In embryonic stem cells derived from a mouse blastocyst, both MOR and KOR promote proliferation and differentiation into neural progenitors through ERK (Kim et al., 2006a). MOR and KOR functionality is found in neural progenitor-derived oligodendrocytes and induces their differentiation via ERK and p38 (Hahn et al., 2010). Activation of opioid receptors blocks the differentiation of both astrocytes and neurons in retinoic acid-induced neural progenitors (Hahn et al., 2010). It is noteworthy that ERK signaling drives the inhibition of astrogenesis, whereas p38 drives the inhibition of neurogenesis.
In early human gestation, both DOR and MOR subtypes are expressed strongly in the SVZ during gestational weeks 11 to 16, but decrease by week 20. DORs and MORs are expressed in multipotential stem cells, newly differentiated neurons, and developing glial cells. However, migrating neurons express negligible levels of either subtype (Tripathi et al., 2008). There is robust expression of KORs in highly enriched (>90% nestin-positive) human fetal brain-derived neural progenitors, and receptor stimulation increases proliferation and migration. These effects are partially blocked by the KOR antagonist, nor-binaltorphimine (Sheng et al., 2007). These many contrasts between these studies on proliferation and differentiation may be due to different mechanisms operating in opioid-mediated neurogenesis versus gliogenesis or may be due to species variations.
In the dentate gyrus of the adult rat hippocampus, all three opioid receptor subtypes have been found both at the mRNA and protein levels (Delfs et al., 1994; Mansour et al., 1995a,b). MOR is highly expressed in the brainstem and spinal cord, regulating functions related to these brain regions such as addiction, alertness, and nociception (Arvidsson et al., 1995; Uhl et al., 1999). However, MORs are also well known to regulate hippocampal function (Morris and Johnston, 1995; Terman et al., 2000; Guo et al., 2005). MORs are expressed on interneurons and primary cell types in all hippocampal regions (Meibach and Maayani, 1980; Arvidsson et al., 1995; Svoboda et al., 1999; Drake and Milner, 2002), including the granule cell layer (GCL) of the dentate gyrus.
The role of opioids in proliferation and differentiation in the adult hippocampus has been complicated and conflicting. Opiates decrease adult SGZ proliferation by inhibiting progenitor proliferation, maturation, and survival (Eisch et al., 2000; Kahn et al., 2005; Arguello et al., 2008) and by altering the progenitor cell cycle (Mandyam et al., 2004, 2007; Arguello et al., 2008). Long-term morphine exposure decreases SGZ neurogenesis by inhibiting dividing cells (particularly those in S phase) and inhibiting progenitor cell progression to a more mature neuronal stage (Arguello et al., 2009). MOR-KO mice exhibit normal proliferation and differentiation but enhanced survival of new neurons and more granule cells (Harburg et al., 2007). In contrast, incubation with the opioid receptor antagonist naloxone reduces proliferation in cultured adult rat hippocampal progenitors with a coincident increase in the differentiation of neurons, but a decrease in astrogliogenesis and oligodendrogenesis is observed (Persson et al., 2003). These inconsistencies are potentially due to the involvement of multiple opioid receptor subtypes in the regulation of neurogenesis and gliogenesis and to differential effects of opioid antagonists on hippocampal progenitor proliferation (Persson et al., 2004).
Opioid receptors have also been shown to decrease proliferation in the SVZ, most studies concentrating on early postnatal animals. Opioid agonists consistently decrease SVZ proliferation in an age-dependent manner. In the postnatal rat SVZ, morphine or β-endorphins, ligands known to interact with opioid receptors in nociception (Bartolome et al., 1994), decrease proliferation in early development but show no effect on older animals (Kornblum et al., 1987; Lorber et al., 1990). In addition, β-endorphin antagonism by a sulfated fragment of cholecystokinin inhibits DNA synthesis (Bartolome et al., 1994). In addition, DNA synthesis in the P11 forebrain is decreased in response to synthetic enkephalin, an opioid receptor agonist (Vértes et al., 1982).
Endogenous opioids modulate adult neurogenesis and gliogenesis in the SVZ by inhibiting cell proliferation as observed in songbirds (Khurshid et al., 2010). These data support an inhibitory mode of action for opioid ligands on cellular proliferation in the SVZ of postnatal mammals. In contrast, studies in the adult rat find that short-term morphine treatment increases [3H]thymidine uptake in the SVZ (Messing et al., 1979; Miller et al., 1982). Cellular proliferation, however, could not be blocked by naltrexone, an opioid receptor antagonist, suggesting that a nonopioid receptor-mediated process may be involved.
J. Peptide Hormones
Vasoactive intestinal polypeptide (VIP) is a peptide neurotransmitter released by GABAergic interneurons in the dentate gyrus. VIP and its receptors (VPAC1 and VPAC2) are expressed in developing and adult dentate gyrus, and VIP blockade causes impairments in associative learning abilities (Gozes et al., 1995).
VIP is known to promote embryonic neurogenesis. It shortens the cell cycle of embryonic neuroepithelial cells (Gressens et al., 1998) and promotes neuronal differentiation of embryonic hippocampal neurons in culture (Blondel et al., 2000). VPAC2 receptor activation shifts the fate of symmetrically dividing neural progenitors toward a nestin-only phenotype, without changing rates of proliferation. In contrast, selective VPAC1 receptor stimulation moves neural progenitor fate toward granule cell neurogenesis (Zaben et al., 2009). Adult Vipr2(−/−) mice exhibit reduced progenitor survival and SGZ neurogenesis, demonstrating a trophic role for VPAC2 receptors in vivo. Type 2 nestin-positive precursors in vivo are also reduced, consistent with a role for VPAC2 in maintaining this cell population (Zaben et al., 2009). This seminal work provides substantial evidence for differential modification of neurogenesis by the VPAC subtypes.
Pituitary adenylate cyclase-activating polypeptide (PACAP) is a 38-amino acid C-terminally α-amidated neuropeptide that was originally isolated from the hypothalamus on the basis of its ability to stimulate adenylyl cyclase activity in rat anterior pituitary cells (Miyata et al., 1989). Alternative processing of the PACAP precursor can generate a 27-amino acid α-amidated peptide (PACAP27) that possesses the same biological activity as the 38-residue peptide (Miyata et al., 1990). PACAP exhibits 68% sequence identity with VIP. The activities of PACAP are mediated through PAC1 receptors (PAC1) that have a much lower affinity for VIP. However, VPAC1 and VPAC2 receptors recognize both PACAP and VIP with high affinity (Vaudry et al., 2000).
During embryonic development, PACAP is expressed in the germinal neuroepithelia and possibly embryonic stem cells. PACAP interacts with sonic hedgehog to reduce proliferation of granule cell precursors (Nicot et al., 2002), increase neurite outgrowth (Gonzalez et al., 1997), and promote survival and differentiation, processes that are required for the development of the cerebellum (Falluel-Morel et al., 2007). PACAP and PAC1 are highly expressed and colocalized in neural progenitors of the mouse cortex at embryonic day 14.5, and induce proliferation via Gq-mediated, but not Gs-mediated, phospholipase C (PLC)/inositol trisphosphate-dependent signaling. These progenitors differentiate mostly into astrocytes (Nishimoto et al., 2007). PACAP also increases DNA synthesis in oligodendrocyte progenitors isolated from neonatal rat brain but delays their maturation (Lee et al., 2001).
PAC1 is expressed in both the SVZ and SGZ of the adult mouse brain and is thought to promote adult neurogenesis. Cultured NSCs/progenitors isolated from the lateral ventricle wall of adult mice express PAC1 and proliferate in vitro in response to two PAC1 agonists, PACAP and maxadilan, but not VIP, at physiologic concentrations. This indicates that PAC1 is a mediator of NSC/progenitor proliferation (Mercer et al., 2004; Ohta et al., 2006). In vitro characterization studies showed that PACAP is capable of inducing NSCs to form multipotent neurospheres that can self-renew and differentiate into both neurons and glia (Mercer et al., 2004) through a PKA-dependent pathway (Ohta et al., 2006). In vivo intracerebroventricular infusion of PACAP increases proliferation in both the SVZ and SGZ (Mercer et al., 2004; Ohta et al., 2006). Recent studies using adult PACAP(−/−) mice indicate decreased survival of newly divided cells in the SGZ, but no difference in proliferation or differentiation compared with control mice under normal or enriched conditions (Ago et al., 2011). In addition, PACAP effectively attenuates apoptosis in the spinal cord of injured rats (Chen and Tzeng, 2005).
VIP is a member of the secretin/VIP/glucagon peptide family and is involved in the positive regulation of embryonic neurogenesis. Secretin (−/−) mice display a decreased number of BrdU-labeled neurons and a dramatic increase in neural progenitor cell apoptosis in in the SGZ during the early postnatal period (Jukkola et al., 2011).
In the adult, VIP also promotes neurogenesis. Adult secretin(−/−) mice have a reduced dentate gyrus volume, decreased long-term potentiation, and impaired spatial learning ability (Jukkola et al., 2011). Taken together, these results suggest a protective and proliferative role in the adult brain for the secretin/VIP/PACAP peptide family that regulates SGZ neurogenesis.
K. Neuropeptide Y
Neuropeptide Y (NPY) is a 36-amino acid peptide that belongs to a family of peptides that includes pancreatic polypeptide and peptide YY. NPY is secreted by the hypothalamus and is widely distributed in the central and peripheral nervous systems. The peptide exerts its biological effects through five GPCRs (Y1–Y5), four of which are functional in humans (Y1, Y2, Y4, Y5).
NPY is an important neuromodulator that regulates mood, endocrine function, blood pressure, nociception, appetite, body weight regulation, and cognition (Michel et al., 1998). Subtypes Y1 and Y5 have known roles in the stimulation of feeding, whereas Y2 and Y4 seem to have roles in appetite inhibition and satiety (Kamiji and Inui, 2007; MacNeil and Douglas, 2007). NPY receptors are also known to be neuroprotective (Xapelli et al., 2006). The Y2 and Y5 receptors were shown to be protective in kainate-induced excitotoxicity in neuronal cultures, as well as in vivo after intrahippocampal kainate injection (Smiałowska et al., 2009). Y2R agonists also show neuroprotective activity in the ischemic middle cerebral artery occlusion model (Smiałowska et al., 2009).
In humans, NPY immunoreactivity is present in all five layers of the cortex of the inferior parietal lobe and the white matter under this cortex (Krivokuća et al., 2010). The dentate gyrus also shows particularly high NPY-like immunoreactivity (Dumont et al., 1992). Particular neuronal cell types that are immunoreactive for NPY include the Cajal-Retzius, known to be involved in embryonic neurogenesis (Bielle et al., 2005) and Alzheimer's disease (Baloyannis, 2005).
NPY increases cell proliferation in the rat SVZ through an ERK pathway but does not affect the self-renewal of NSCs (Thiriet et al., 2011). NPY is also involved in ATP-induced neuroproliferation in adult mouse olfactory epithelium (Jia and Hegg, 2010). Using KO mice in vivo or cell cultures with specific NPY receptor agonists and antagonists, the neuroproliferative and neuronal differentiating effects of NPY in the SVZ were found to be mediated by the Y1 receptor subtype through ERK-mediated signaling (Hansel et al., 2001; Howell et al., 2003, 2005; Agasse et al., 2008; Decressac et al., 2009). Mice with a targeted deletion of NPY contain half as many dividing olfactory neuronal precursor cells as do controls and develop significantly fewer olfactory neurons by adulthood (Hansel et al., 2001).
NPY promotes SGZ neurogenesis as well, and this effect is also thought to be mediated by the Y1 receptor subtype through ERK signaling (Hansel et al., 2001; Howell et al., 2003, 2005; Agasse et al., 2008; Decressac et al., 2009). Intracerebroventricular administration of NPY increases SGZ cell proliferation and promotes neuronal differentiation in adult mice through the Y1, but not Y2, receptor (Decressac et al., 2011). Furthermore, NSCs/progenitors from the postnatal hippocampus are sensitive to the combination of NPY and FGF2, which significantly shortens the cell cycle time of nestin-positive NSCs more than either factor alone, enhancing rates of proliferation (Rodrigo et al., 2010).
NPY cells express other neurotransmitters, such as nitric-oxide synthase, which influence progenitor cell proliferation, migration, and neurite outgrowth (Reif et al., 2004; Chen et al., 2005). Within the hippocampus, there is an increasing appreciation of the role of GABAergic interneurons as a mechanism for excitation-neurogenesis coupling (Deisseroth and Malenka, 2005). Many in vivo manipulations that affect electrical activity influence neuron production from neural stem/progenitor cells in the dentate gyrus (for review, see Lehmann et al., 2005). GABAergic excitation is also known to promote neuronal differentiation in these cells (Tozuka et al., 2005). NPY is coreleased with GABA by GABAergic interneurons under high-frequency firing conditions and induces proliferation of nestin-positive NSCs cultured from the whole hippocampus (Howell et al., 2003).
Seizures are known to induce NPY expression (Cardoso et al., 2010). NPY then partly mediates seizure-induced NSC proliferation in the SGZ in vivo (Howell et al., 2007). Furthermore, Y1-deficient mice show decreased seizure-induced proliferation in the SGZ, as well as in the subcallosal zone, where seizures induce glial proliferation (Howell et al., 2007).
L. Purinergic Receptors
Two families of purinergic GPCRs exist: adenosine receptors (or P1 receptors) and P2Y receptors (Fredholm et al., 1994). In humans, there are four adenosine receptors (A1, A2A, A2B, A3), with the A1 and A2A receptors playing important roles in the brain. Adenosine is the endogenous ligand for the adenosine receptors (Fredholm et al., 2001). P2Y receptors are another family of purinergic GPCRs that is stimulated by nucleotides such as ATP, ADP, UTP, UDP, and UDP-glucose. To date, 12 P2Y receptors have been cloned in humans: P2Y1–2, 4–6, 8–14 (Abbracchio et al., 2006). The gaps in the numbering of the P2Y subtypes are due to previous incorrect classification.
Adenosine modulates neuronal activity presynaptically by inhibiting or facilitating transmitter release and postsynaptically by affecting the action of other neurotransmitters. It acts nonsynaptically by hyperpolarizing or depolarizing neurons (Ribeiro et al., 2002). Adenosine has been shown to play protective roles in the hippocampus, which is highly sensitive to ischemia and hypoxia (Sebastião et al., 2001).
The amount of information on the neurogenic effects of the P1 adenosine receptor is very limited. Cells isolated from the mouse fetal midbrain increase neuron formation when adenosine is applied (Delic and Zimmermann, 2010). However, there are no studies exploring neurogenic potential of P1 adenosine receptors in adult cell or animal models.
ATP-dependent purinergic signaling through the P2Y1 receptor has been associated with developmental neurogenesis (Weissman et al., 2004), as radial glial signaling required for embryonic neurogenesis involves P2Y1 ATP receptors. Embryonic NPCs express P2Y purinergic receptors and release ATP themselves to mobilize intracellular calcium and increase progenitor proliferation. Furthermore, P2Y receptor antagonists suppress proliferation and instead promote differentiation into neurons and glia in vitro, whereas subsequent removal of purinergic inhibition restores progenitor cell expansion (Lin et al., 2007). ATP, UTP, and adenosine-5′-O-(2-thiodiphosphate), in conjunction with EGF, induce converging signals that cause proliferation and then guide adult neural progenitor migration through the formation of stress fibers, actin cytoskeleton, AKT, and focal adhesion kinase in cultured adult murine NSCs (Grimm et al., 2009; Grimm et al., 2010).
In human NSCs, ATP and P2Y receptors have been identified as mitogens for v-myc immortalized neural progenitor cells through calcium release and PI3K (Ryu et al., 2003). Activation of P2Y induces DNA synthesis in glial cells (Neary and Zhu, 1994; Milenkovic et al., 2003) as well as radial glia (Uckermann et al., 2002). NTPDase, which degrades active ATP, is selectively localized to the adult rat SVZ along with P2Y1 and P2Y4 receptors (Lin et al., 2007). In the adult mouse SVZ, type B cells and residual radial glia cells in the hippocampus selectively express NTPDase2 (Shukla et al., 2005; Mishra et al., 2006; Langer et al., 2007). In addition, neurospheres cultured from the adult mouse SVZ express NTDPase2, P2Y1, and P2Y2 receptors. P2Y1 and P2Y2 receptor agonism augments proliferation in the presence of growth factors, whereas neurospheres isolated from P2Y1-deficient mice show reduced proliferation (Mishra et al., 2006). In another study using the selective P2Y1 receptor antagonist 2′-deoxy-N6-methyladenosine 3′,5′-bisphosphate (MRS 2179), primary neurospheres from the adult mouse SVZ demonstrated A2A and P2Y1 receptor-mediated inhibition of neurosphere generation and proliferation (Stafford et al., 2007). However, MRS 2179 may be only ∼11-fold selective for the P2Y1 over the P2X1 receptor ligand-gated ion channels (Brown et al., 2000).
Previous studies have shown that expression of P2Y receptors in astrocytes is altered when connexin43, a gap junction protein, is down-regulated (Suadicani et al., 2003) and that the growth rate of Cx43-null astrocytes is decreased compared with that of WT cells (Dermietzel et al., 2000). In embryonic neurospheres isolated from connexin 43-deficient mice, the concomitant decrease in P2Y1 receptors was found to regulate the decrease in proliferation and migration as a result of this gap-junction channel loss (Scemes et al., 2003). These results are consistent with an in vitro model of CNS differentiation in which neural progenitor cells were found to be coupled by Cx43 channels, and blockade of gap junctional communication strongly reduced proliferation and differentiation of neural progenitors (Duval et al., 2002).
M. Serotonin
Serotonin, or 5-hydroxytryptamine (5-HT), plays a major role in the CNS and is an important regulator of adult neurogenesis (Banasr et al., 2004; Malberg, 2004; Warner-Schmidt and Duman, 2006). Reduced 5-HT function resulting in impaired adult neurogenesis has been hypothesized to be involved in major depression (Duman et al., 2000; Jacobs et al., 2000; Kempermann, 2002). Serotonergic neurons innervate the entire CNS, originating from cells in the raphe nuclei (Lorez and Richards, 1982) and the reticular formation of the brain stem. At least 15 subtypes of serotonin receptors exist, including 5-HT1–5 and 5-HT7. Within each receptor type, there are subcategories of receptors. Except for the 5-HT3 receptor, all of the 5-HT receptors are GPCRs. The roles of the 5-HT1A and 5-HT2A/2C receptor subtypes in adult neurogenesis are the most understood.
Selective serotonin reuptake inhibitors (SSRIs) increase 5-HT levels by blocking its reuptake at the presynaptic serotonergic nerve terminal. Long-term SSRI treatment, which is commonly used to decrease symptoms of depression and other mood disorders, has been shown to increase cell proliferation in both the adult rat hippocampus in vivo and in neural cultures in vitro (Malberg et al., 2000; Lee et al., 2001; Manev et al., 2001). The SSRI-induced proliferative effect seems to be mediated predominantly through activation of 5-HT1A receptors (Santarelli et al., 2003), in a postsynaptic effect (Huang and Herbert, 2005). Administration of a partial 5-HT1A agonist will also increase adult neurogenesis (Grabiec et al., 2009) through a similar mechanism. In contrast, 5-HT1A receptor blockade reduces cell proliferation in the dentate gyrus (Radley and Jacobs, 2002). However, no change in cell proliferation is seen in 5-HT1A receptor-KO mice (Santarelli et al., 2003).
Lesions to serotonergic inputs result in decreased adult neurogenesis in both the SGZ and SVZ (Brezun and Daszuta, 1999), which can be functionally restored by the transplantation of serotonergic tissue to the lesion site (Brezun and Daszuta, 2000a,b). Serotonin depletion also reduces survival and proliferation in cultured neurospheres derived from adult mouse hippocampus (Benninghoff et al., 2010).
Evidence suggests that 5-HT2 receptors are also involved in regulating neurogenesis, especially in the SVZ. In addition to 5-HT1A receptor activation, 5-HT2 receptor agonists enhance cell proliferation in the SZV (Banasr et al., 2004). Oddly, 5-HT2A/2C antagonists have also been found to increase adult neurogenesis in the SVZ (Wang et al., 2004; Green et al., 2006). A previous study found that fluoxetine enhances the division of early progenitor (TAP) cells but not stem-like cells (Encinas et al., 2006). It has been discovered that serotonin may exert differential effects on neural stem/progenitor cells (Soumier et al., 2010) via opposing actions mediated through different 5-HT receptors during different stages.
In the adult hippocampus, long-term 5-HT1A receptor activation and 5-HT2A blockade increased proliferation, whereas long-term 5-HT2 receptor stimulation reduced proliferation. Long-term 5-HT2 receptor stimulation increased survival and enhanced maturation (Klempin et al., 2010). Other studies have shown that long-term 5-HT2A/2C blockade increases the proliferation of progenitor cells in the adult hippocampus (Jha et al., 2008). It is noteworthy that 5-HT4 receptor agonists have been discovered to increase adult neurogenesis in a much shorter time frame than SSRIs, possibly by increasing serotonergic drive and enhancing cAMP-responsive element-binding protein phosphorylation (Lucas et al., 2007). The overall effect of the various 5-HT receptor-mediated actions of serotonin is increased adult neurogenesis. Because of its critical role in depression and other mood disorders, how and to what extent serotonin affects adult neurogenesis is an important question that warrants extensive further investigation.
N. Wnt
Wnt proteins bind to receptors of the atypical GPCR frizzled and low-density lysoprotein-related protein5/6 families on the cell surface. To date, at least 17 human genes have been cloned for Wnt signaling glycoproteins: Wnt1, -2, -2B, -3, -3A, -4, -5A, -5B, -6, -7A, -7B, -8A, -8B, -9A, -9B, -10A, -10B, -11, and -16 (Katoh and Katoh, 2005). Wnt binding to its receptors induces the nuclear translocation of β-catenin that acts as a transcriptional cofactor of lymphoid enhancer binding factor 1 and modifies gene transcription (canonical Wnt pathway). Wnt can also signal in noncanonical pathways independent of β-catenin, leading to small G-protein activation and cytoskeletal changes or Ca2+ signaling with activation of heterotrimeric G-proteins, causing various cell responses (Force et al., 2007).
Wnt receptors are implicated in adult hippocampal neurogenesis through the regulation of proliferation, migration, and differentiation (Zhou et al., 2004; Lie et al., 2005; Yu et al., 2006; Wexler et al., 2009). The Wnt 3 receptor is expressed and the β-catenin pathway is active in the hippocampal SGZ. Overexpression of Wnt3 is sufficient to increase adult neurogenesis in the SGZ in vitro and in vivo. In contrast, blockade of Wnt signaling abolishes neurogenesis almost completely in vivo (Lie et al., 2005). Low-density lysoprotein-related protein 6 mutant mice exhibit reduced production and proliferation of dentate granule neurons and abnormalities in the radial glial scaffolding (Zhou et al., 2004). In a series of in vitro studies using adult mouse neurospheres, both Wnt3 and Wnt5a promoted neurogenesis, but only Wnt5a-mediated neurogenesis was blocked by PKC inhibition (Yu et al., 2006).
O. Intracellular Signals (Phospholipase C-β1, Phospholipase A2, Phosphodiesterase-4D)
Direct downstream second messengers and effectors that are normally associated with GPCR signaling may be able to mediate adult neurogenesis independently of any GPCR system. These signals include arachidonic acid, diacyl glycerol, inositol 1,4,5-triphosphate, and cAMP.
Enzymes that metabolize arachidonic acid, such as 5-lipoxygenase, cyclooxygenase-2, and prostaglandin E2, have been reported to increase proliferation of neuronal precursors in the adult SGZ when infused into the hippocampus (Uchida et al., 2002). Cyclooxygenase-2-deficient mice show lower numbers of proliferating cells, which are induced to proliferate in the postischemic SGZ (Sasaki et al., 2004). Several phospholipase A2 isoforms are neurotrophic for cerebellar granule neurons (Arioka et al., 2005) or regulate neurite outgrowth and survival in culture (Smalheiser et al., 1996; Forlenza et al., 2007; Masuda et al., 2008).
PLC is an enzyme that cleaves phosphatidylinositol 4,5-bisphosphate into diacyl glycerol and inositol 1,4,5-trisphosphate. PLC-β1(−/−) mice show increased adult hippocampal neurogenesis, increased adult granule cell density and survival, and stable proliferation rates (Manning et al., 2012). These mice also display aberrant migration of mature granule neurons within the GCL in adulthood, with excessive adult-generated mature neurons residing in the middle and outer GCL.
Mice deficient in phosphodiesterase-4D (PDE4D), an enzyme that catalyzes the hydrolysis of cAMP, display memory enhancement in radial arm maze, water maze, and object recognition tests, along with increased hippocampal neurogenesis. Microinfusion of lentiviral vectors that contained microRNAs targeting long-form PDE4D isoforms into bilateral dentate gyri of the mouse hippocampus down-regulate PDE4D4 and PDE4D5, enhance memory, and raise hippocampal neurogenesis (Li et al., 2011). Long-term PDE4 inhibition by rolipram treatment also increases adult neurogenesis, as evidenced by enhanced proliferation and survival of BrdU-positive neurons in the hippocampal dentate gyrus (Sasaki et al., 2007; Li et al., 2009). This suggests that PDE4D normally acts to repress hippocampal neurogenesis.
VII. Implications of Adult Neurogenesis in Pathological Conditions
Adult neurogenesis is altered in brain injuries and several neurological diseases and disorders, including traumatic brain injury and ischemic stroke; neurodegenerative diseases such as Alzheimer's, Huntington's, and Parkinson's diseases (Abdipranoto et al., 2008); demyelinating diseases such as multiple sclerosis; epilepsy and seizures; and psychiatric disorders such as depression and schizophrenia. A detailed review on neurogenesis in pathological states is found in Kempermann (2011a).
A. Adult Neurogenesis in Brain Injury
Traumatic brain injury (TBI) occurs when sudden trauma causes damage to the brain. It is the most common form of acquired brain injury in children, young adults, and older people. Recovery after TBI depends on the balance between neuronal injury and neuroregeneration. TBI typically causes neuronal loss and neurological deficits, especially in hippocampus-dependent cognitive functions. Accumulating evidence from animal studies of both focal and diffuse TBI suggests that neurogenesis is increased after TBI in both the SGZ of the dentate gyrus (Kernie et al., 2001; Yu et al., 2008; Zheng et al., 2011) and the SVZ of the lateral ventricles (Bye et al., 2011). However, it remains to be demonstrated conclusively whether neurogenesis contributes to recovery after TBI.
Stroke is the leading cause of disability and death in humans and is the result of ischemia, blockage, or hemorrhage to the brain. Numerous studies indicate that focal or global ischemia potently stimulate neurogenesis in adult rodents (Liu et al., 1998; Jin et al., 2001; Kee et al., 2001) and monkeys (Tonchev et al., 2005; Koketsu et al., 2006). In contrast, subarachnoid hemorrhage has been shown to be associated with a decrease in cell proliferation in the SGZ and SVZ (Mino et al., 2003). Although functional neurogenesis has been demonstrated in the hippocampus (van Praag et al., 2002) and newly born neurons in the SVZ have been shown to migrate to the damaged cortex (Arvidsson et al., 2002), the role of ischemia-induced neurogenesis in recovery after an ischemic insult remains unclear.
B. Adult Neurogenesis in Neurodegenerative Diseases
1. Alzheimer's Disease.
Alzheimer's disease (AD) is a progressive neurodegenerative disease of the CNS and the most common cause of dementia in older people. AD is characterized by the accumulation of plaques and tangles in the brain, particularly in the hippocampus. The plaques contain insoluble deposits of amyloid-β-peptide, whereas the neurofibrillary tangles are composed of aggregates of the hyperphosphorylated microtubule-associated protein τ. The cognitive deficits observed in AD correlate most closely with the τ pathology (Selkoe, 2003; Zhang et al., 2012). The role of neurogenesis in AD is unclear and confusing. Increased hippocampal neurogenesis is observed in some animal models of AD, including the Sorl1 KO mouse (Rohe et al., 2008) and mutated APP23 mouse model (Mirochnic et al., 2009). DCX is expressed in neurons in the SGZ of the dentate gyrus; consistent with this, postmortem samples taken from patients with AD possess more DCX-positive cells, suggesting that adult neurogenesis is increased (Jin et al., 2004a). In contrast, adult neurogenesis is reduced in double transgenic mice for mutated APP and presenilin-1 (Niidome et al., 2008) and triple transgenic mice for APP, presilin-1, and τ (Rodríguez et al., 2008). Furthermore, hippocampal volume is reduced in people with AD (Roh et al., 2011). Surprisingly, regulation of neurogenesis seems to remain intact in these models of AD. Reduced overall cholinergic signaling is a well known hallmark of AD, and loss of cholinergic drive has been shown to reduce adult neurogenesis in both the hippocampus and olfactory bulb (Cooper-Kuhn et al., 2004). Conversely, treatment with cholinergic drugs, which increase cholinergic signaling, has been shown to stimulate neurogenesis (Mohapel et al., 2005; Kotani et al., 2006).
2. Huntington's Disease.
Huntington's disease (HD) is a progressive neurodegenerative genetic disorder of the CNS. Initially, HD affects muscle coordination; eventually, however, it leads to cognitive decline and dementia. It is caused by an autosomal-dominant mutation on the Huntingtin (HTT) gene, resulting in an expansion of a CAG triplet repeat stretch and alteration in the subsequent Htt protein. The altered Htt protein causes gradual pathological changes to the brain, giving rise to a wide spectrum of movement, cognitive, and psychiatric signs and symptoms. In genetic animal models of HD, such as the R6 transgenic mouse, adult neurogenesis is reduced (Lazic et al., 2004; Gil et al., 2005). In contrast, increased cell proliferation and neurogenesis is observed in postmortem HD human brain tissue (Curtis et al., 2003).
3. Parkinson's Disease.
Parkinson's disease is a progressive degenerative disorder of the CNS that affects movement. It results from the death of dopaminergic neurons in the substantial nigra. Signs and symptoms of PD include tremor, bradykinesia, muscle rigidity, impaired posture, speech changes, and eventually dementia. PD is characterized by the accumulation of α-synuclein aggregates in the form of Lewy bodies. There is little evidence supporting active neurogenesis in the adult substantia nigra. Furthermore, there does not seem to be reactive neurogenesis in either the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) or 6-OHDA animal models of PD. One study reported increased BrdU labeling in the substantia nigra after MPTP treatment as well as data supporting neurogenesis in the normal adult substantial nigra (Zhao et al., 2003), whereas another study found no new neurons in the substantia nigra of rodents injected with 6-OHDA (Frielingsdorf et al., 2004). However, after dopaminergic lesions with 6-OHDA or MPTP, the number of intrinsic tyrosine hydroxylase-positive (dopamine, NE) neurons increases in the striatum, and there is a transient elevation of adult neurogenesis in the hippocampus (Winner et al., 2009; Park and Enikolopov, 2010). Recent studies suggest that upon injury, reactive astrocytes promote adult DA neurogenesis through the Wnt1/β-catenin signaling pathway (L'Episcopo et al., 2011, 2012). In contrast, decreased neurogenesis has been shown in both the SGZ and SVZ in PD (Höglinger et al., 2004).
4. Amyotrophic Lateral Sclerosis.
Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disease characterized by rapidly progressive muscle weakness, disability, and death. It is caused by the degeneration of lower motor neurons located in the spinal cord and the upper motor/cortical neurons in the brainstem and cortex. In mice carrying a mutation of superoxide dismutase 1, a mutation found only in rare familial patients with ALS, increased progenitor cell proliferation, migration, and neurogenesis has been reported (Chi et al., 2006). Currently, there is no effective treatment for ALS.
Adult neurogenesis and neural plasticity is modulated in several neurodegenerative diseases. It is noteworthy that genes often involved in neurodegenerative diseases such as PD (α-synuclein), AD (presenilin-1, τ), and HD (huntingtin) also seem to play important roles in regulating adult neurogenesis (for review, see Winner et al., 2011).
C. Adult Neurogenesis in Demyelinating Disease
Multiple sclerosis (MS) is an inflammatory autoimmune disease characterized by demyelination and axonal scarring. This leads to less effective communication between nerve cells in the brain and spinal cord. MS occurs most often in young adults and women (Compston and Coles, 2008) and is often associated with periods of worsening (relapsing) and improvement (remitting), as some remyelination takes place. Unfortunately, oligodendrocytes, the cells responsible for the making and maintaining of myelin in the CNS, cannot completely rebuild the sheath, because MS destroys oligodendrocytes. In the adult brain, oligodendrocytes arise from a subset of neuron-glial antigen 2-expressing perinatal OPCs (Menn et al., 2006). Normally, few new oligodendrocytes are generated in the adult brain (McCarthy and Leblond, 1988; Ehninger and Kempermann, 2003); however, after demyelination injury, some neuron-glial antigen 2 cells will give rise to oligodendrocytes (Redwine and Armstrong, 1998; Nait-Oumesmar et al., 1999; Levine et al., 2001). Furthermore, increased neurogenesis is observed in long-term lesions of MS (Chang et al., 2008). As the disease advances, the remyelination becomes less effective (Wolswijk, 1998), and neurologic function progressively deteriorates. Patients with MS typically live 5 to 10 years fewer than those without the disease. A recent report suggests that activation of gliogenesis in the SVZ occurs in MS and that SVZ-derived early glial progenitors may give rise to OPCs (Nait-Oumesmar et al., 2007). These early glial progenitors could be a potential therapeutic target for strategies to promoting myelin repair in MS.
D. Adult Neurogenesis in Epilepsy and Seizures
Epilepsy, characterized by recurrent seizures, is the third most common neurological disorder after stroke and AD. Epileptic seizures result from abnormal synchronous neuronal activity in the brain (Fisher et al., 2005). Temporal lobe epilepsy (TLE) is the most common form of epilepsy in adult humans and is often associated with hippocampal lesions. In animal models of epilepsy, adult neurogenesis is profoundly enhanced in both the SGZ and SVZ (Parent et al., 1998; Ferland et al., 2002; Jessberger et al., 2007a). Precursor cell proliferation and adult neurogenesis may increase as much as 10-fold after short-term seizures. The magnitude of the increase depends on the severity of the seizures; mild seizures cause a smaller increase than severe (Yang et al., 2008a). Evidence of increased cell proliferation has also been found in tissue from children with hippocampal sclerosis and seizures (Takei et al., 2007). However, chronic TLE is associated with decreased adult hippocampal neurogenesis (Hattiangady et al., 2004). In addition, the seizure-induced increase in neurogenesis seems to decline with age and can become completely absent in old age (Rao et al., 2008; Shetty et al., 2011). In fact, short-term seizures in old age may lead to marked neuronal loss, a greater chance of developing chronic TLE, and diminished cognitive function (Hattiangady et al., 2011).
Seizures, in addition to enhancing neurogenesis, seem to abnormally affect neuronal development, polarity, migration, and integration (Jessberger et al., 2007b; Kraev et al., 2009). Ectopic neurons are found in the hilus and cornu ammonis 3 regions in rodent model of epilepsy (Parent et al., 1999) as well as in tissue from humans with chronic TLE (Thom et al., 2002). Many of these ectopic neurons seem to have incorrectly integrated into the network, because they can be activated by cornu ammonis 3 pyramidal cell firing but not by granule cells (Scharfman et al., 2000; McCloskey et al., 2006). It has been hypothesized that this seizure-induced aberrant neurogenesis may actually contribute to the development of epilepsy (Parent, 2007; Hattiangady et al., 2008; Zhao and Overstreet, 2008). Clearly, the role of neurogenesis in seizures and epilepsy merits further investigation.
E. Adult Neurogenesis in Psychiatric Disorders
1. Major Depression.
Major depressive disorder (MDD) is a psychiatric disorder characterized by generalized low mood, low self-esteem, and a loss of interest or pleasure in activities that are usually enjoyable. It is the most prevalent mental health disorder in the United States. The lifetime risk of MDD is almost 25%, and lifetime risk of suicide among untreated patients is nearly 20%; approximately 10% of Americans suffer depression in a given year (Kessler and Wang, 2009).
The idea that depression is due to impaired adult neurogenesis was first proposed in 2000 (Jacobs et al., 2000). A plethora of experimental data has been found that supports the neurogenesis hypothesis of depression; however, an equally compelling amount of evidence exists against the theory (for review, see Kempermann et al., 2008; Samuels and Hen, 2011). The hippocampi of patients with chronic MDD are often smaller (Sheline et al., 1996; Sheline, 2000). Stress and cortisol levels, which are often elevated in people with MDD, reduce adult hippocampal neurogenesis (Malberg and Duman, 2003). Antidepressants such as the SSRIs increase cell proliferation in the adult hippocampus (Malberg et al., 2000; Santarelli et al., 2003; Encinas et al., 2006; Pinnock et al., 2009). In addition, the negative effects of stress on adult hippocampal neurogenesis are reversed by the SSRI fluoxetine (Malberg and Duman, 2003). Conversely, the antidepressive action of fluoxetine is removed when adult hippocampal neurogenesis is abolished (Santarelli et al., 2003). More recent studies suggest that these proneurogenic effects are species-dependent (Holick et al., 2008) as well as dependent on the particular antidepressant used (Surget et al., 2008). It is now thought that fluoxetine's antipressant effect possesses both neurogenesis-dependent and -independent components (David et al., 2009). It is noteworthy that several proneurogenic compounds have been found to produce a positive response in tests of anxiety or depression, including FGF2 (Perez et al., 2009) and Notch1 (Guo et al., 2009). Environmental enrichment and exercise, which increase adult hippocampal neurogenesis, are known to improve mood (Van Praag et al., 1999; Veena et al., 2009). Current knowledge suggests that impaired adult neurogenesis plays an important role in MDD, although MDD is not a purely neurogenesis-based disorder (Kempermann, 2011a).
2. Schizophrenia.
Schizophrenia is a severe chronic disease affecting higher cognitive functions. It is characterized by progressive disintegration of thought processes and emotional responsiveness and often manifests in some combination of hallucinations, paranoia, and disordered speech, thinking, and behavior. The onset of schizophrenia typically occurs in young adults, with lifetime prevalence of schizophrenia at almost 1%. The ability to function normally progressively deteriorates, resulting in significant social and occupational dysfunction. As a result of increased health problems and a very high suicide rate of nearly 5%, patients with schizophrenia typically live 12 to 15 years fewer than unaffected people (van Os and Kapur, 2009).
Decreased adult neurogenesis has been hypothesized to be a contributory factor in schizophrenia (for review, see Kempermann et al., 2008). Hippocampal cell proliferation is decreased in postmortem tissue of patients with schizophrenia (Reif et al., 2006). Furthermore, whole-brain and hippocampal volume is smaller in people with schizophrenia (Steen et al., 2006).
The cause of schizophrenia is unknown, but several genes linked to schizophrenia have also been associated with neurogenesis (Le Strat et al., 2009). The Disc 1 (disrupted in schizophrenia) gene was found in a Scottish family with a high incidence of schizophrenia as well as major depression (Millar et al., 2000). It is highly expressed in neurogenic areas and seems to play an important role in the development and integration of new neurons (Duan et al., 2007) through the AKT-mammalian target of rapamycin pathway (Kim et al., 2009). Disc 1 has also been shown to regulate neurogenesis through comodulation of Wnt-glycogen synthase kinase 3β/β-catenin signaling (Mao et al., 2009). Mutations of Disc 1 or glycogen synthase kinase 3β impair adult neurogenesis (Eom and Jope, 2009).
VIII. Adult Neural Stem Cell Therapy in the Central Nervous System
Adult stem cells possess two major defining properties, the ability to self-renew and the ability to differentiate into multiple specialized cell types. In the mammalian CNS, adult NSCs are able to generate new functional neurons and glia. Although active adult neurogenesis seems to be mostly restricted to the neurogenic SGZ and SVZ regions in the brain, it is exhibited in other areas of the CNS (for review, see Kempermann, 2011b). Adult neurogenesis plays an active and important role in maintaining normal homeostatic processes and contributes to the plasticity of the CNS (for review, see Lledo et al., 2006; Ming and Song, 2011).
The role of adult neurogenesis in neurological disorders and diseases is still poorly characterized and is currently a major research focus. Adult neurogenesis may help to repair the CNS in response to brain injury or disease. In general, acute brain damage tends to transiently increased cell proliferation, whereas chronic neuropathology eventually results in decreased adult neurogenesis. In rodents, NSCs are involved in replacing neurons dying from direct injury (Magavi et al., 2000) or ischemic stroke (Arvidsson et al., 2002; Nakatomi et al., 2002). Furthermore, it seems that cell proliferation and neurogenesis initially increase at sites of damage in many seizure models (Bengzon et al., 1997; Parent et al., 1998; Jessberger et al., 2005) and neurodegenerative diseases such AD (Jin et al., 2004a; Van Kampen and Eckman, 2006; López-Toledano and Shelanski, 2007) and HD (Curtis et al., 2003). Increased neuroplasticity, a known function of adult neurogenesis, is also observed in many human neurodegenerative disorders (Kempermann, 2011a). NSC-based therapies hold great promise for treating many of the aforementioned neurological disorders. The two major strategies currently used in NSC-based therapy include transplantation of exogenous neural stem/progenitor cells and stimulation of endogenous neural stem/progenitor cells (Taupin, 2006).
Transplantation of fetal neuronal precursor cells has shown some success in animal models of spinal cord injury (McDonald et al., 1999), TBI (Riess et al., 2002), PD (Bjorklund et al., 2002), and spinal muscular atrophy (Corti et al., 2010). However, similar neurotransplantation therapies in humans with PD or HD have yielded mixed results. Whereas some patients with PD exhibited remarkable improvements (Wenning et al., 1997), others showed no benefits (Olanow et al., 2003), and some suffered additional side effects (Hagell et al., 2002). Clinical trials in patients with HD have also encountered problems, as some neurotransplants displayed HD-like neuronal degeneration (Cicchetti et al., 2009), whereas one patient developed mass lesions (Keene et al., 2009).
Adult neuronal precursor cells are also used for transplantation therapy. Adult spinal cord stem cells transplanted locally into the adult dentate gyrus have been shown to generate new neurons in rats (Shihabuddin et al., 2000). Furthermore, neurospheres derived from adult neural stem/progenitor cells administered either intravenously or intrathecally resulted in remyelination and functional recovery in a mouse model of MS (Pluchino et al., 2003).
There are many challenges and technical issues that must be overcome before neurotransplantation of adult or embryonic derived stem/progenitor cells can proceed much further in humans (for review, see Feng and Gao, 2012). Challenges associated with transplanting these cells into the CNS include improper delivery and migration, mass effect and cell death, cell manufacturing problems, tumor formation, and adverse immune system interactions (Chiu and Rao, 2011).
Stimulation of endogenous neural stem/progenitor cells affords another potential strategy for treating neurological disorders and diseases. Although adult neurogenesis is mostly restricted to the neurogenic SGZ and SVZ regions in the brain, neural stem/progenitor cells seem to reside throughout the CNS (Kempermann, 2011b). The recruitment and activation of these endogenous cells would represent a strategy to promote regeneration and repair of the damaged or diseased CNS.
There are multiple methods to stimulate endogenous neurogenesis for the replacement of lost cells that have not yet been explored. For example, stimulating NSCs in the SVZ to replicate and migrate to the sites of damage and/or degeneration via the rostral migratory stream and possibly the corpus callosum fiber tract (Osman et al., 2011). In theory, this could be accomplished either by direct or indirect stimulation of the appropriate neural stem/progenitor cells. Direct stimulation could include the application of an agent that directly binds to and stimulates the NSC (perhaps through a GPCR-mediated system). Indirect stimulation could involve an agent that produces its effect by causing and/or increasing the release of some other endogenous agent (possibly by inhibiting an enzyme or a neurotransmitter reuptake mechanism), which in turn directly stimulates the NSC. Proneurogenic endogenous therapeutic strategies are currently being pursued by a number of pharmaceutical corporations and biotech companies.
IX. Therapeutic Potential of G-Protein-Coupled Receptor-Based Neural Stem Cell Strategies
Numerous compounds have been found to be proneurogenic. Antidepressant SSRIs are probably the best known, most studied, and some of the few proneurogenic agents used clinically (serotonin, NE, and dopamine reuptake inhibitors, or a combination, are all probably proneurogenic). SSRIs are indirect proneurogenic compounds that produce their effect by increasing 5-HT levels, which in turn stimulate neurogenesis via the activation of 5-HT receptors. In addition to the 5-HT receptors and other GPCR neurotransmitter systems mentioned in this review (section VI), a variety of other agents can enhance adult neurogenesis, including nitric oxide donors, acetylcholinesterase inhibitors, statins, N-methyl-d-aspartate receptor blockers, anti-inflammatory agents, and sex hormones (for review, see Jang et al., 2008; Kempermann, 2011c). The major disadvantage with most of these compunds, including the SSRIs, is a lack of specificity for their proneurogenic effects.
Research efforts have recently been undertaken to discover more specific targets and compounds for proneurogenic drug development. Some of these efforts have focused on identifying GPCRs that selectively stimulate NCS activity and/or proliferation of NPCs and differentiation into particular neuronal cell types. These attributes would prove valuable for both stimulation of in vivo endogenous adult neurogenesis and in vitro culture of cells for exogenous transplantation therapy. To date, most research on GPCR-based NSC therapies has focused on stimulating endogenous adult neurogenesis in vivo, either alone or in combination with another agent. Using a combination of in vitro screening, genomics, and bioassays, followed by in vivo neurogenesis and functional behavioral assays, several small-molecule compounds have recently been discovered that interact with GPCRs to stimulate NSC activity. GPCR-mediated proneurogenic drug strategies currently under investigation and/or clinical development include 5-HT receptor agents, α1A-AR compounds, CB2 receptor agonists, DOR peptides, melatonergic agents, mGlu2/3 receptor antagonists, and S1P or LPA receptor compounds. Long-term α1A-AR stimulation improves learning and memory, reduces anxiety and depression, and increases lifespan in mice (Doze et al., 2009, 2011; Perez and Doze, 2011). Early preclinical and phase 1 trials with 5-HT receptor agents and mGlu2/3 antagonists have shown promise. The major neurological disorders targeted by GPCR-based NSC therapies thus far include psychiatric disorders, such as anxiety and depression, and select neurodegenerative diseases, including AD, HD, PD, and ALS.
There is a pressing need for better treatments for many neurological disorders, particularly the currently incurable CNS diseases. Because adult neurogenesis seems to be modifiable, stimulation of endogenous neurogenic processes, and/or the use of transplanted exogenous NSCs, has been postulated as a possible therapeutic strategy for several neurological disorders. Hopes have been raised by the prospect of NSC therapy. However, despite intense research efforts, successful NSC therapy has remained elusive. Developing effective strategies for isolating, enriching, and propagating NSCs for transplantation has proven problematic. Furthermore, identifying the molecules and cellular mechanisms required for the proper integration of transplanted stem cells into the injured brain has been difficult. As the processes that regulate adult neurogenesis are more fully understood, it is expected that the dream of functional NSC therapy will be reached. As demonstrated in this review, considerable information of the role of GPCRs in adult neurogenesis has been obtained. Using this knowledge, GPCR-based NSC therapeutic strategies may soon be realized.
X. Concluding Remarks and Future Directions
Many tissues of the body can repair and self-renew, largely because of stem cells and the various mechanisms that regulate their behavior. The concept that the brain does not regenerate was one of the most recent bastions of science shattered. Thus, the science of NSCs is a relatively new field. Although advances are slowly being made, there is a huge potential to ameliorate neurological diseases and counteract the aging process. The major weakness in the scientific evidence for adult neurogenesis is the lack of human research. Although evidence in rats and mice is encouraging, its application to human diseases is not assured. In particular, clinical trials will need to be performed to determine whether the production of new neurons can play therapeutic roles. Given the inherent difficulties of measuring neurogenesis in humans, the identification of molecular correlates and the signal transduction pathways involved in adult neurogenesis may prove to be the best course for future research.
The addition of new neurons to the existing circuitry represents a prominent form of structural plasticity and may contribute to specific brain functions in the adult brain. Research over the past decade has established a close relationship between adult neurogenesis and neuronal activity. Various environmental cues, exercise, learning and cognition-related stimuli, and emotional activities can influence neurogenic activity. With the addition of pharmaceuticals as suggested through GPCR ligands, we may be able to enhance our cognitive functions as well as treat neurological diseases.
Acknowledgments
This work was supported in part by the National Science Foundation [Grant 0347259]; the National Institutes of Health National Center for Research Resources [Grant P20-RR016471]; the National Heart, Lung and Blood Institute [Grant R01-HL098279]; and a Grant in Aid from the American Heart Association, Great Rivers Affiliate. We thank Katie Collette, Haley Amoth, and Elizabeth Sandquist for assistance in editing the manuscript.
Authorship Contributions
Wrote or contributed to the writing of the manuscript: Doze and Perez.
Footnotes
This article is available online at http://pharmrev.aspetjournals.org.
↵1 Abbreviations:
- 5-HT
- 5-hydroxytryptamine
- 6-OHDA
- 6-hydroxydopamine
- AD
- Alzheimer's disease
- ALS
- amyotrophic lateral sclerosis
- AR
- adrenergic receptor
- BrdU
- bromodeoxyuridine
- CB
- cannabinoid
- CBD
- cannabidiol
- CNS
- central nervous system
- CNTF
- ciliary neurotrophic factor
- CSF
- cerebrospinal fluid
- DCX
- doublecortin
- DOR
- δ-opioid receptor
- DSP-4
- N-(2-chloroethyl)-N-ethyl-2-bromo benzylamine hydrochloride
- EGF
- epidermal growth factor
- ERK
- extracellular signal-regulated kinase
- FGF
- fibroblast growth factor
- GCL
- granule cell layer
- GFAP
- glial fibrillary acidic protein
- GFP
- green fluorescent protein
- GPCR
- G-protein-coupled receptor
- h
- human
- KO
- knockout
- KOR
- κ-opioid receptor
- LPA
- lysophosphatidic acid
- MDD
- major depressive disorder
- mGluR
- metabotropic glutamate receptor
- MGS0039
- (1R,2R,3R,5R,6R)-2-amino-3-(3,4-dichlorobenzyloxy)-6-fluorobicyclo[3.1.0]hexane-2,6-dicarboxylic acid
- miRNA
- microRNA
- MOR
- μ-opioid receptor
- MPTP
- 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine
- MRS 2179
- 2′-deoxy-N6-methyladenosine 3′,5′-bisphosphate
- MS
- multiple sclerosis
- NCAM
- neural cell adhesion molecule
- NE
- norepinephrine
- NPY
- neuropeptide Y
- NTPDase
- nucleoside triphosphate diphosphatase
- OPC
- oligodendrocyte progenitor cell
- PACAP
- pituitary adenylate cyclase-activating polypeptide
- PD
- Parkinson's disease
- PDE
- phosphodiesterase
- PI3K
- phosphoinositide 3-kinase
- PKC
- protein kinase C
- PLC
- phospholipase C
- S1P
- sphingosine-1-phosphate
- SDF-1
- stromal cell-derived factor 1
- SGZ
- subgranular zone
- SKF 38393
- 2,3,4,5-tetrahydro-7,8-dihydroxy-1-phenyl-1H-3-benzazepine
- SNC80
- (+)-4-[(αR)-α-(2S,5R)-4-allyl-2,5-dimethyl-1-piperazinyl)-3-methoxybenzyl]-N,N-diethylbenzamide
- Sox2
- Sex-determining region Y-box 2
- SSRI
- selective serotonin reuptake inhibitor
- SVZ
- subventricular zone
- TAP
- transient amplifying progenitor
- TBI
- traumatic brain injury
- THC
- Δ9-tetrahydrocannabinol
- TLE
- temporal lobe epilepsy
- U50488H
- trans-(±)-3,4-dichloro-N-methyl-N-[2-(1-pyrrolidinyl)cyclohexyl]benzeneacetamide
- VIP
- vasoactive intestinal polypeptide
- VPAC
- vasoactive intestinal polypeptide receptor
- VPC32183
- (S)-phosphoric acid mono-{2-octadec-9-enoylamino-3-[4-(pyridin-2-ylmethoxy)-phenyl]-propyl} ester
- WT
- wild type.
- © 2012 by The American Society for Pharmacology and Experimental Therapeutics
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
In this issue





{kind=link}
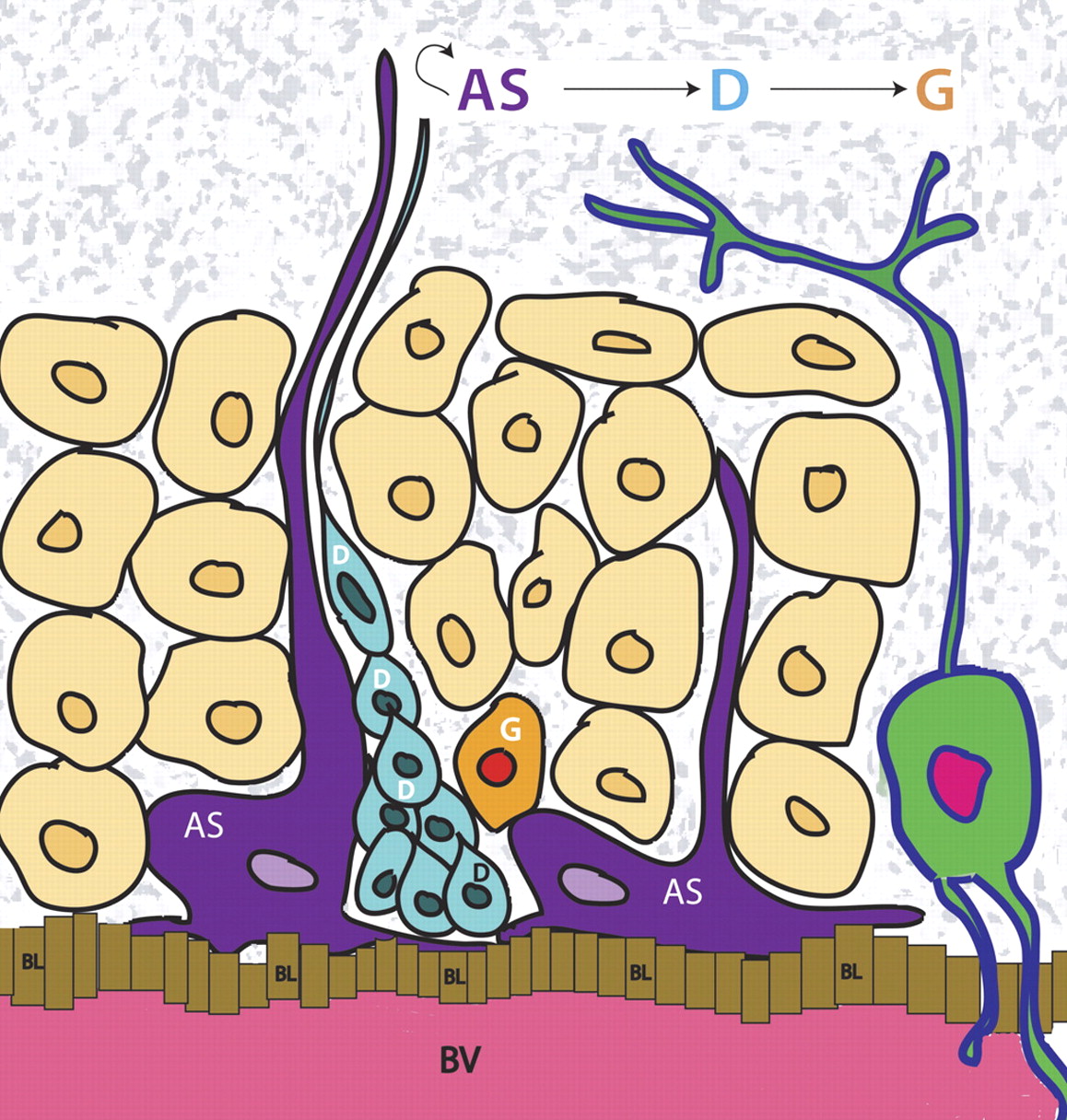{kind=link}
Jump to section
- Article
- Abstract
- I. Introduction
- II. General Features of Neural Stem Cells and Progenitors
- III. Adult versus Embryonic Neurogenesis
- IV. Methods for Analyzing Adult Neurogenesis
- V. Regulation of Adult Neurogenesis in the Brain Vascular Niche and Choroid Plexus
- VI. G-Protein-Coupled Receptor Regulation of Adult Neurogenesis
- VII. Implications of Adult Neurogenesis in Pathological Conditions
- VIII. Adult Neural Stem Cell Therapy in the Central Nervous System
- IX. Therapeutic Potential of G-Protein-Coupled Receptor-Based Neural Stem Cell Strategies
- X. Concluding Remarks and Future Directions
- Acknowledgments
- Authorship Contributions
- Footnotes
- References
- Figures & Data
- Info & Metrics
- eLetters