Article Text

Abstract
Background: Fragile X syndrome (FXS) is the most common single gene inherited form of mental retardation, with behaviours at the extreme of the autistic spectrum. Subjects with FXS and fragile X mental retardation gene knock out (Fmr1 KO) mice, an animal model for FXS, have been shown to exhibit defects in dendritic spine maturation that may underlie cognitive and behavioural abnormalities in FXS. Minocycline is a tetracycline analogue that has been used in clinical trials for stroke, multiple sclerosis and several neurodegenerative conditions.
Methods: We evaluated the effects of minocycline on dendritic spine development in the hippocampus of young Fmr1 KO mice, and in primary cultures of hippocampal neurons isolated from those mice. Cognitive effects of minocycline in young WT and Fmr1 KO mice were also evaluated using established behavioural tests for general cognition, activity and anxiety.
Results: Our studies demonstrate that minocycline promotes dendritic spine maturation both in cultures and in vivo. The beneficial effects of minocycline on dendritic spine morphology are also accompanied by changes in the behavioural performance of 3-week-old Fmr1 KO mice. Minocycline treated Fmr1 KO mice show less anxiety in the elevated plus maze and more strategic exploratory behaviour in the Y maze as compared to untreated Fmr1 KO mice. Our data suggest that these effects of minocycline may relate to its inhibitory action on MMP-9 expression and activity, which are higher in the hippocampus of Fmr1 KO mice.
Conclusion: These findings establish minocycline as a promising therapeutic for the treatment of fragile X mental retardation.
Statistics from Altmetric.com
Fragile X syndrome (FXS) is the most common single gene inherited form of mental retardation and autism. Subjects with FXS often show visual–spatial memory problems, attention deficits, developmental delays, hyperactivity,1 and childhood seizures.2 Amplification of CGG repeats in the non-coding 5′ region of the fragile X mental retardation (FMR1) gene leads to gene silencing and FMR1 protein (FMRP) deficiency. In mice, Fmr1 mRNA and FMRP are dynamically regulated during early postnatal development.3 As FMRP localises to post-synaptic sites and can bind to ∼4% of brain mRNA transcripts,4 it could regulate protein translation and/or mRNA localisation of key synaptic components. Indeed, FMRP has been shown to regulate the translation of post-synaptic molecules in dendritic spines.3–6 Fmr1 knockout (KO) mice display abnormal socialisation, signs of obsessive/compulsive behaviour, and are highly susceptible to audiogenic seizures.7
Abnormal dendritic spine development is a common feature in the brains of subjects with FXS, Down and Rett syndromes.8 9 Dendritic spines are specialised surface protrusions on the dendrites that receive the majority of excitatory synapses, and changes in their morphology impact synaptic plasticity and long term memory.10–12 Initially, dendritic spines develop as long, thin filopodia-like protrusions that make contact with pre-synaptic targets and then mature into short, stubby or mushroom shaped structures.13–16 Mice deficient for FMRP (Fmr1 KO) show a higher proportion of long, thin filopodia than wild-type (WT) littermates.17 18 Dendritic spine abnormalities in Fmr1 KO mice are most pronounced during development,18 19 but persist into adulthood.20 Molecular links between FMRP deficiency and a preponderance of immature dendritic spines are unresolved, but may involve group 1 metabotropic glutamate receptor (mGluR) hyperactivity21 or alterations in protein regulators of dendritic spine maturation.22
Here we show that minocycline treatment induces mature dendritic spine morphology in the hippocampus of young Fmr1 KO mice, and in primary cultures of hippocampal neurons isolated from those mice. Further, the beneficial effects of minocycline on dendritic spine maturation in the hippocampus of Fmr1 KO mice were accompanied by behavioural changes in young mice. Minocycline is a tetracycline analogue that can inhibit matrix metalloproteinase-9 (MMP-9) and reduce inflammation in the central nervous system (CNS). Recent clinical trials have tested minocycline as a possible treatment for stroke, multiple sclerosis (MS), several neurodegenerative conditions, and most recently autism. Our findings suggest that the beneficial effects of minocycline in young Fmr1 KO mice may relate to its inhibitory action on MMP-9 expression and activity, which is higher in FMRP deficient hippocampal neurons as compared to wild-type (WT) neurons. Moreover, our results suggest that minocycline may provide beneficial effects to patients with FXS.
MATERIALS AND METHODS
Mice
The FVB/NJ-Fmr1tm1Cgr (Fmr1 KO) and FVB/NJ controls (WT) were obtained from the Jackson Laboratories. Mice were maintained in an AAALAC accredited facility under 12 h light/dark cycles. All mouse studies were done within National Institutes of Health and institutional animal care and use committee guidelines.
Minocycline administration in vivo
Minocycline was administered to newborn mice (WT and Fmr1 KO mice) by adding it to the mother’s drinking water at 30 mg/kg/day every day for 7–28 days. This method has been previously reported to result in high concentrations of minocycline in the breast milk of lactating mothers. Minocycline administration through drinking water has been shown to decrease significantly MMP-9 expression and activity in the adult brain due to its ability to cross the blood–brain barrier.23
Behavioural analysis with the elevated plus and Y maze
Seven to 16 male mice from each of four groups (minocycline treated and non-treated, Fmr1 KO and WT mice) were tested for anxiety in an elevated plus maze (see supplemental material).24 Mice were also tested for spontaneous alternation behaviour (SAB) using a Y maze (see supplemental material).25 Mice on the FVB background have been reported to undergo retinal degeneration at a young age involving the loss of most rods and cones except those thought to project to the suprachiasmatic nucleus for circadian rhythms.26 Photoreceptor degeneration begins at 2 weeks of age and progresses for up to 2 months. At 2 months of age the mice begin to show a significant decrease in visual acuity.27 We tested mice at 3 weeks before significant degeneration had occurred. Minocycline treated mice received the drug through their lactating mother’s drinking water for 4 weeks, until weaning when minocycline treatment was stopped. For elevated plus and Y maze, one way ANOVAs were used. Following ANOVA, post hoc pair-by-pair differences between groups were resolved with the least significant difference (LSD) using Dunnett’s method and Hsu’s Multiple Comparison with the Best treatment (MCB).
Slices
Coronal 300 μm thick sections were prepared from the brains (prefixed in 4% paraformaldehyde to preserve morphology) of P7 WT and Fmr1 KO mice treated with minocycline or control water (total: 12 mice, three per condition). Developing hippocampal neurons were labelled in slices with the red fluorescent membrane dye DiI using a biolistic particle mediated gene transfer method (gene gun) as previously described (see supplemental material).28 Morphometric analysis of dendritic spines and quantification of the numbers and lengths of dendritic spines, as well as sizes of dendritic spine heads, were performed as previously described (see supplemental material).29 30
Hippocampal neuron cultures
Cultures of hippocampal neurons were prepared from embryonic day 15 (E15) or E16 mouse embryos of WT and Fmr1 KO mice as previously described (supplemental material).30
MMP-9 and minocycline treatments in vitro
Recombinant mouse MMP-9 (CC069, Chemicon-Millipore, Temecula, California, USA) was activated with 1 mM APMA (A9563, Sigma, St Louis, Missouri, USA) in 100 mM Tris-HCl buffer (pH 7.6) containing 3 μM ZnCl2, 10 mM CaCl2 and 0.02% Brij 35 (activation buffer) at 37°C for 2 h. Hippocampal cultures were treated with 100 ng/ml MMP-9 in Neurobasal medium for 1 h under 5% CO2/10% O2 atmosphere at 37°C. Minocycline (155718, MP Biomedicals, Solon, Ohio, USA) was added to hippocampal cultures at 20 μM for 17 h. The treatments were performed in conditioned medium under 5% CO2/10% O2 atmosphere at 37°C.
Immunostaining and image analysis were performed as previously described (supplemental material).31
Biochemical analysis
Gelatin gel zymography and western immunoblotting were performed as previously described31 32 with minor modifications (supplemental material).
RESULTS
Minocycline promotes the formation of mature dendritic spines in cultured hippocampal neurons
We first examined the effects of minocycline on dendritic spine morphology in cultured hippocampal neurons (fig 1A). Although minocycline treatment had no significant effects on average dendritic spine length and density, it promoted the formation of larger dendritic spine heads in 2-week-old cultures (fig 1B–D). WT neurons treated with minocycline showed more rapid dendritic spine maturation, with a higher proportion of mushroom shaped spines (fig 1E). Importantly, minocycline was well tolerated by hippocampal neurons, with no neurotoxicity at concentrations as high as 20 μg/ml (data not shown). These results establish that minocycline promotes the formation of mature mushroom shaped spines in cultured hippocampal neurons.
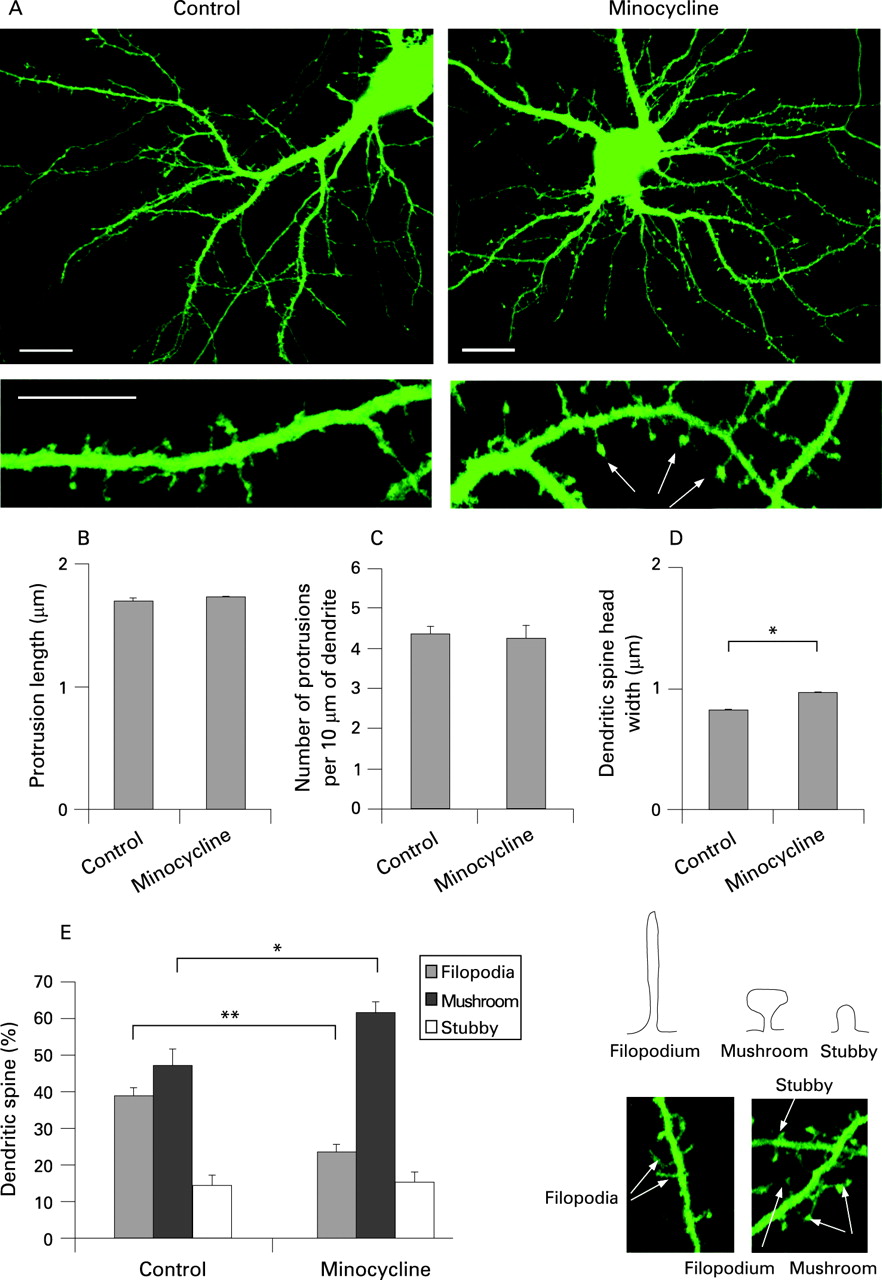
Minocycline treatment induces the maturation of dendritic spines in Fmr1 KO hippocampal neurons
We observed, as reported by others,19 that hippocampal neurons from Fmr1 KO mice exhibit immature dendritic spine profiles with longer spines, a lower density of mushroom shaped spines with large heads and excessive filopodia in comparison to WT controls at 15 DIV (fig 2). As minocycline accelerated dendritic spine development in WT neurons, we tested if it could do the same in Fmr1 KO neurons and diminish the immature dendritic spine profile. Minocycline treatment of Fmr1 KO hippocampal neurons significantly increased the proportion of mushroom shaped spines in 15 DIV cultures to the levels that were similar to age matched WT controls (fig 2). Interestingly, minocycline treatment did not change dendritic spine length or the number of dendritic spines in Fmr1 KO neurons, but did reduce the density of immature filopodia-like dendritic spines with a corresponding increase of mushroom spines with large heads. These findings suggest that minocycline reverses the immature dendritic spine phenotype of Fmr1 KO neurons in vitro by transforming immature dendritic spines into mature spines.

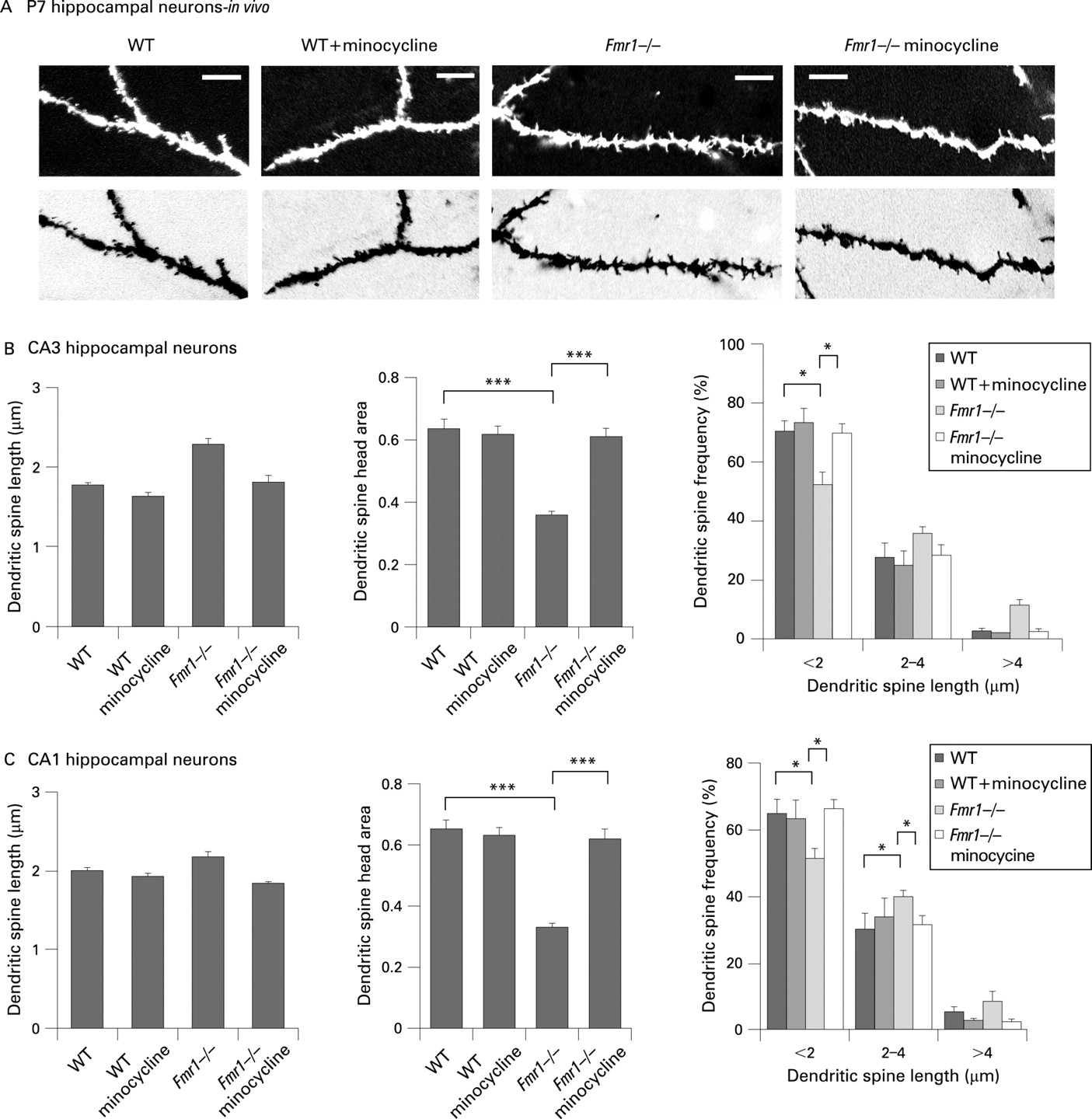
Minocycline treatment of Fmr1 KO mice reduces dendritic spine abnormalities in the hippocampus
As many properties of neurons are different in vitro and in vivo, we also performed in vivo analysis of dendritic spine development in the hippocampus of WT and Fmr1 KO mice. Morphometric analysis of dendritic spines showed an increase in average dendritic spine length in the hippocampus of postnatal day 7 (P7) Fmr1 KO mice, as compared to WT control mice (fig 3), which corroborated our in vitro studies. Fmr1 KO neurons also exhibited dendritic spines with smaller heads, an increased proportion of longer spines with length 2.00–4.00 μm and >4.00 μm, and a significant decrease in the density of shorter spines with length <2.0 μm, in both CA3 and CA1 areas as compared to control WT neurons. Further, we investigated the effects of minocycline administration on dendritic spine development in the hippocampus of newborn Fmr1 KO mice. Minocycline was administered to newborn mice (WT and Fmr1 KO mice) by adding it to the mother’s drinking water at 30 mg/kg/day for 7 days. Minocycline administration increased the proportion of short dendritic spines with larger heads in both CA3 and CA1 areas of the hippocampus of P7 Fmr1 KO mice, resulting in dendritic spine profiles that were very similar to WT control mice (fig 3). These studies conclusively demonstrate that minocycline treatment can induce dendritic spine maturation in the hippocampus of Fmr1 KO mice, correcting aberrant dendritic spine development caused by FMRP deficiency.

Higher anxiety in Fmr1 KO mice is reduced by minocycline treatment
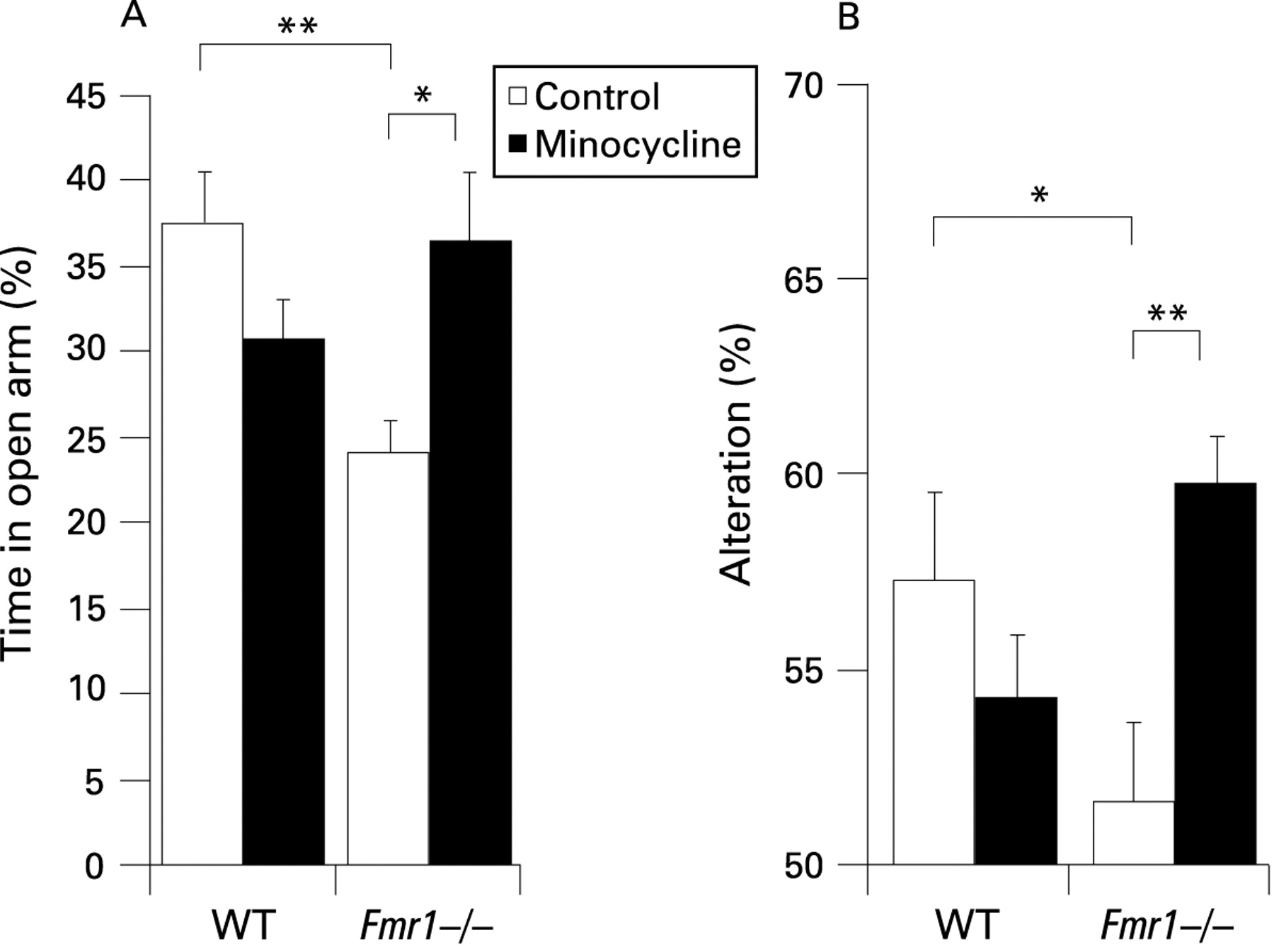
Next, we examined whether the effects of minocycline on dendritic spine morphology in the hippocampus of Fmr1 KO mice was accompanied by changes in behavioural performance. Young Fmr1 KO and WT mice were tested for anxiety in an elevated plus maze. The maze consisted of four radial arms, two open and two closed by tall walls. Anxious mice naturally avoid open spaces and spend more time in the closed arms, but less anxious mice spend more time exploring the open arms. We digitally recorded all mice in the plus maze, and used time spent in the open arms as an indicator of anxiety. At 3 weeks of age Fmr1 KO mice spent significantly (p = 0.0059) less time in the open arms than WT mice (fig 4A). However, there was no significant difference between WT and Fmr1 KO mice that were nursed by mothers given minocycline in their drinking water. Minocycline was administered to newborn mice (WT and Fmr1 KO mice) by adding it to the mother’s drinking water at 30 mg/kg/day for 21 days. This increase of time spent in the open arms by minocycline treated Fmr1 KO mice was significantly higher (p = 0.0124) than untreated Fmr1 KO mice, indicating they were less anxious.24 These findings demonstrate that Fmr1 KO mice have behavioural deficits compared with WT mice at 3 weeks of age, which are significantly improved in Fmr1 KO mice treated with minocycline.
Minocycline improves the behavioural performance of Fmr1 KO mice in a hippocampus dependent spontaneous alternation task
Although the Morris and radial arm water maze tasks are often used for the evaluation of hippocampus dependent behaviour,33 young mice (3 weeks) panic and would be in peril of drowning. Therefore, we evaluated hippocampus dependent memory using a passive Y maze where the spontaneous alternation behaviour of mice could be observed.27 The maze consisted of three equally spaced arms with tall walls. Mice were allowed to explore the Y maze for 6 min, and digitally recorded from above with the tester out of the room. If mice exited arm A and entered arm B, an alternation was scored if the next arm entered was C. If they returned to arm A (A>B>A) then they did not alternate, but if they went into arm C (A>B>C) then an alternation had occurred. In strictly random arm entries there would be 50% alternation. However, alternations above 50% indicate some memory of which arm had been most recently explored. All arm entries in a 5 min trial were scored by a blinded observer from digital videos. Minocycline treated Fmr1 KO mice made significantly (p = 0.0021) more alternations than untreated Fmr1 KO mice (fig 4B). Notably, there were no significant differences between untreated and minocycline treated WT mice. Minocycline treatment had no effect on total arm entries (supplemental fig 1).
These findings demonstrate that early treatment with minocycline (<4 weeks) has benefits on the behaviour of Fmr1 KO mice, with minimal effects on the behavioural performance of WT mice.
Minocycline treatment lowers MMP-9 in the hippocampus of Fmr1 KO mice
To determine if minocycline affects MMP-9 expression and activity in vivo we examined the levels of pro and active forms in protein lysates isolated from the hippocampus of untreated or minocycline treated Fmr1 KO mice by immunoblotting with an anti-MMP-9 antibody. Concentrations of active MMP-9 were higher in P7 hippocampal lysates from Fmr1 KO mice, as compared to WT controls (fig 5A). Gel zymography showed significantly higher MMP-9 gelatinase activity in lysates from Fmr1 KO hippocampus (fig 5B). Further, minocycline treatment significantly reduced levels of MMP-9 in Fmr1 KO hippocampus (fig 5A). MMP-9 gelatinase activity was also significantly lower in the hippocampus of minocycline treated Fmr1 KO mice (fig 5C). Our findings suggest that the beneficial effects of minocycline on dendritic spine maturation in the hippocampus of Fmr1 KO mice may relate to its inhibitory actions on MMP-9 expression and activity.

MMP-9 shifts hippocampal neurons toward an immature dendritic spine profile
We recently reported that MMP-7 treatment of WT hippocampal neurons induces a dramatic redistribution of F-actin and alters dendritic spine morphology, transforming dendritic spines from mushroom-like into filopodia-like protrusions.31 Although MMP-7 is not normally expressed in hippocampal neurons, it can process neuronally expressed pro-MMP-9, which is produced in the developing hippocampus, into the fully active form. Here we found that MMP-9 also induced immature dendritic spine profiles in 15 DIV hippocampal neuron cultures, although the effects were less profound than with MMP-7. There were fewer short dendritic spines (<2.0 μm) with large heads (0.5–1.0 μm) and more long dendritic spines (2.0–4.0 μm) with small heads (<0.5 μm) in MMP-9 treated cultures as compared to controls (fig 6C,E). MMP-9 treatment also significantly reduced the average size of dendritic spine heads (fig 6D). Analysis of dendritic spine morphology also revealed that MMP-9 treated neurons exhibited more filopodia and fewer mushroom shaped dendritic spines (fig 6F). In summary, while minocycline promoted dendritic spine maturation, MMP-9 induced immature dendritic spine morphology.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
We have found that minocycline reverses abnormal behaviours in Fmr1 KO mice, an animal model for FXS. We observed that 3-week-old Fmr1 KO mice spent less time in the open arms of an elevated plus maze than WT controls, indicating higher anxiety. Minocycline treatment reduced anxiety in Fmr1 KO mice such that they spent the same amount of time in open arms as WT mice. Similar results were also seen in the Y maze, a spontaneous alternation test, where 3-week-old Fmr1 KO mice showed no exploration strategy. Remarkably, early minocycline treatment improved the alternation behaviour of Fmr1 KO mice to levels that were at least as good as WT controls, often better. The beneficial effects of minocycline on the behaviour of Fmr1 KO mice was significant at 3 weeks of age.
Our analyses of minocycline effects on synapse development demonstrate that this drug promotes dendritic spine maturation in vivo and in vitro. Significantly, this effect is sufficient to overcome deficits in dendritic spine maturation that occur in Fmr1 KO mice. Abnormal synapse and dendritic spine development is a common feature in FXS and may represent the anatomical and physiological basis for cognitive and behavioural abnormalities associated with this disorder.34 Several studies have established strong correlations between the size of dendritic spine heads and synaptic strength, wherein filopodia-like spines with smaller heads have fewer AMPA receptors (AMPARs)35 and are less stable than mushroom-like spines with larger heads.36 The development of dendritic spines is abnormal in the brains of subjects with FXS, Down and Rett syndromes, as they show a preponderance of long, thin filopodia-like spines at ages when control subjects have more mature, mushroom shaped dendritic spines.8 9 Our findings demonstrate the beneficial effects of minocycline in FMRP deficient neurons and in the brains of Fmr1 KO mice by transforming long, thin spines into mature mushroom shaped spines.
Mechanisms responsible for the prevalence of immature dendritic spines in Fmr1 KO mice remain unclear, but may be related to metabotropic glutamate receptor (mGluR) hyperactivity. An mGluR theory of FXS posits that FMRP normally suppresses protein synthesis that attenuates mGluR dependent long term depression (LTD). FMRP deficiency leads to group 1 mGluR hyperactivity, resulting in enhanced LTD.37 This theory is supported by the observation that exaggerated activation of group 1 mGluR and increased mGluR induced LTD are present in the hippocampus of Fmr1 KO mice.38 Pharmacological activation of group 1 mGluRs with the specific agonist DHPG also induces dendritic spine elongation,21 producing a morphology that is similar to what we have observed in hippocampal neurons treated with MMP-7 or MMP-9. Moreover, mGluR5 inhibition with the specific antagonist MPEP suppresses two behavioural phenotypes of Fmr1 KO mice: (1) an elevated susceptibility to audiogenic seizures (AGS); and (2) an increased tendency to move to the centre of an open field.7 Our studies demonstrate that DHPG also induces MMP-9 expression and activation in cultured hippocampal neurons (supplemental fig 2), suggesting that MMP-9 activation could be induced by group 1 mGluR hyperactivity and contribute to abnormal dendritic spine development in FMRP deficient hippocampal neurons.
Minocycline effects on dendritic spine morphology may also be related to its inhibitory actions on MMP-9 expression and activity, which is upregulated in FMRP deficient hippocampal neurons. While minocycline induced dendritic spine maturation in both WT and FMRP deficient hippocampal neurons in vitro, the in vivo effects of minocycline on dendritic spines and behaviour, in wild-type mice, were not significant and much less dramatic than in the Fmr1 KO animals. WT mice express endogenous MMP-9 in the hippocampus that participates in long term potentiation and learning, but the Fmr1 KO mice have higher levels of MMP-9 (fig 5). Although the mechanism responsible for higher MMP-9 expression in Fmr1 KO mice is still unclear, it is possible that high basal levels of MMP-9 activity in the brains of Fmr1 KO mice may interfere with normal physiological responses and induce dendritic spine remodelling. Indeed, our findings indicate that excessive MMP-9 activity disrupts mature dendritic spines in hippocampal neurons (fig 6). Therefore, lowering MMP-9 activity with minocycline may offset these destabilising effects of MMP-9, resulting in the more significant effects on Fmr1 KO neurons than with WT neurons. Consistent with our findings, proteolytically active MMP-9 has been shown to induce slowly emerging synaptic potentiation in the CA1 area of the hippocampus and to occlude tetanically evoked LTP in intact rat brain.39 Our findings indicate that minocycline may also have beneficial effects in neurodevelopmental disorders that exhibit elevated brain concentrations of MMP-9.
MMP-9 could affect dendritic spine morphology by cleaving components of the extracellular matrix (ECM) and/or cell surface proteins that participate in synaptogenesis and dendritic spine maturation.40 For example, MMP-9 mediated cleavage of perineuronal ECM networks could release RGD containing peptides that bind to integrins and facilitate NMDAR activation. We have previously shown that RGD containing peptides can induce rapid dendritic spine elongation in cultured hippocampal neurons, mediated by NMDAR.30 Furthermore, MMP-7 effects on dendritic spines are similar to those of RGD and are also blocked by the NMDAR specific inhibitor MK-801.31 While abnormally high MMP-7 or MMP-9 activities interfere with normal physiological functions and induce dendritic spine remodelling, modest concentrations of MMP-9 have been shown to regulate non-pathological synaptic functions and plasticity in mature hippocampus through an integrin dependent mechanism and NMDAR activation.39 Integrins are known to modulate NMDAR mediated synaptic currents and play an important role in LTP.41 42 Although there is a clear connection between MMP-9 and integrin signalling, other MMP-9 substrates in synapses may also be involved, including pro-BDNF, cadherins and Eph/ephrins.40 Interestingly, while FMRP does not affect the expression levels of BDNF or its high affinity receptor TrkB, BDNF infusion fully restores LTP in slices from Fmr1 KO mice, suggesting that BDNF activity modulation may be a potentially useful therapeutic strategy for FXS.43 Notably, subjects with FXS may also be affected by MMP dysregulation outside the CNS as they display several characteristics of modified ECM regulation including large ears, long faces, hyperextensible joints and extremely soft skin.
In addition to the inhibitory effects of minocycline on MMP-9 expression and activity, minocycline can also inhibit microglial activation by suppressing p38 MAPK activity.44 Although it is possible that the inhibition of microglia proliferation may contribute to the dendritic spine effects of minocycline in vivo, in our hippocampal neuron cultures microglia represented <1% of the cells (not shown), suggesting that minocycline effects on dendritic spine morphology are independent of microglial actions, at least in vitro. Moreover, minocycline has been shown to inhibit apoptosis by downregulating caspase activities and the inducible form of nitric oxide synthetase (iNOS) in a transgenic model of Huntington disease.45 Anti-apoptotic actions of minocycline have also been reported in a mouse model of amyotrophic lateral sclerosis, and in experimental models of retinal cell death and light induced retinal degeneration.46 47 However, it is possible that the minocycline actions on late apoptotic events, such as cytochrome c release and caspase activation, are mediated through the regulation of protein synthesis or by inhibiting MMP expression and activity, which would alter cell–ECM interactions.
It remains to be determined if MMP-9 contributes to abnormal dendritic spine development and behavioural deficits in Fmr1 KO mice, but these findings establish that minocycline can supersede FMRP deficiency to achieve normal dendritic spine profiles in Fmr1 KO mice and improve behavioural performance. These findings establish the beneficial effects of minocycline in the Fmr1 KO mouse model and indicate a high therapeutic potential for this drug, or related compounds, in the treatment of human subjects with FXS.
Acknowledgments
We thank Dr Michael Tranfaglia and members of the Ethell laboratories for helpful discussions and Hani El Shawa for technical help.
REFERENCES
Supplementary materials
web only appendices 46/2/94
Files in this Data Supplement:
Footnotes
Additional material and figures are published online only at http://jmg.bmj.com/content/vol46/issue2
Funding: This work is supported by The FRAXA Research Foundation.
Competing interests: None.