

Abstract
It is increasingly appreciated that executive control processes need to be understood in terms of motivational as well as cognitive mechanisms. The current study examined the impact of performance-contingent reward incentives (monetary bonuses) on neural activity dynamics during cued task-switching performance. Behavioral measures indicated that performance was improved and task-switch costs selectively reduced on incentive trials. Trial-by-trial fluctuations in incentive value were associated with activation in reward-related brain regions (dopaminergic midbrain, paracingulate cortex) and also modulated the dynamics of switch-selective activation in the brain cognitive control network. Within lateral prefrontal cortex (PFC), both additive (inferior frontal junction) and interactive [dorsolateral PFC (DLPFC)] incentive effects were observed. In DLPFC, incentive modulation of activation predicted task-switching behavioral performance, but with hemispherically dissociable effects. Furthermore, in left DLPFC, incentive modulation specifically enhanced task–cue-related activation, and this activation in turn predicted that the trial would be subsequently rewarded (because of optimal performance). The results suggest that motivational incentives have a selective effect on brain regions that subserve cognitive control processes during task-switching and, moreover, that one mechanism of effect might be the enhancement of cue-related task preparation within left DLPFC.
Introduction
Complex human behavior is dependent on the ability to effectively engage cognitive control, a collection of processes enabling the formation, maintenance, and updating of behavioral goal representations, as well as top-down biasing mechanisms that enhance goal-relevant information processing, inhibit goal-irrelevant information, and detect conflicts between them (Desimone and Duncan, 1995; Smith and Jonides, 1999; Monsell and Driver, 2000; Botvinick et al., 2001; Miller and Cohen, 2001; Engle, 2002; Braver and Ruge, 2006; O'Reilly, 2006; Boucher et al., 2007). Cognitive control is thought to be critical for task preparation: the configuration of attention, perception, and action selection systems before task engagement based on contextual information (Monsell and Driver, 2000). Such preparatory processes have been successfully studied using cued task-switching paradigms, in which advance cues convey that one of multiple possible tasks is to be performed on an upcoming ambiguous stimulus (Rogers and Monsell, 1995; Meiran, 2000; Mayr, 2006; Monsell and Mizon, 2006). Previous work in human and animal cognitive neuroscience has implicated the lateral prefrontal cortex (PFC) and parietal cortex as centrally involved in preparatory processing during task-switching (Sohn et al., 2000; Crone et al., 2006; Yeung et al., 2006; Johnston et al., 2007; Ruge and Braver, 2007; Stoet and Snyder, 2007; Rowe et al., 2008; Kamigaki et al., 2009).
An important, but oftentimes implicit, assumption in theoretical models is that cognitive control success, or failure, is dependent on the activation strength of the cued task goal, yet it is still unclear what are the most important factors that modulate goal activation strength. Motivational variables represent a likely candidate for this function, because motivational states are widely thought to regulate behavioral goal salience, the priority of goal pursuit, and criteria governing goal completion (Kruglanski et al., 2002). Nevertheless, only recently have research investigations begun to focus on the neural mechanisms associated with motivation: cognitive control interactions (Pessoa, 2008, 2009). In the current study, we examine how motivation enhances preparatory cognitive control by isolating the effect of performance-contingent reward incentives on cue-related versus target-related processing during task-switching.
Neurophysiological studies in behaving primates have provided explicit evidence for motivational influences on neural mechanisms of cognitive control, by demonstrating that the presence of reward incentives can modulate delay-related activity in lateral PFC neurons during maintenance periods of working memory tasks (Watanabe, 1996; Leon and Shadlen, 1999). Human neuroimaging studies have confirmed this work by demonstrating reward-related modulations of activity in PFC, parietal cortex, and associated neural systems during working memory and executive control task performance (Pochon et al., 2002; Gilbert and Fiez, 2004; Taylor et al., 2004; Small et al., 2005; Krawczyk et al., 2007; Locke and Braver, 2008; Kouneiher et al., 2009). However, the previous studies have not tightly linked changes in neural activity with subsequent motivation-related enhancements in behavioral performance. Moreover, there remains a need to demonstrate that performance-contingent reward incentives enhance cognitive control selectively. Cued task-switching paradigms provide an excellent opportunity to examine such questions, by enabling (1) comparison of incentive effects under switching versus single-task conditions and (2) isolation of preparatory control processes associated with the encoding of advance task cues.
Here, we report results demonstrating that trial-by-trial fluctuations in incentive reward value modulate switch-selective activation in prefrontal and parietal components of the cognitive control network (CCN). Moreover, we demonstrate that, in the left dorsolateral PFC (DLPFC), such reward-related activation increases are preparatory in nature and predict behavioral performance enhancements.
Materials and Methods
Participants.
Sixteen participants (eight male, eight female), aged 18–29 years (mean ± SD, 22 ± 2.3 years), were recruited from Washington University to participate in the study. All participants were right-handed, native English speakers, had corrected-to-normal vision, were free from psychiatric or neurological disorders, and had no contraindications to functional magnetic resonance imaging (fMRI). The Washington University institutional review board approved the study, and all participants provided written consent before participation. Compensation was provided for participation, in terms of base amounts for the behavioral practice session ($10/h) and fMRI imaging session ($25/h), plus an additional bonus of $10 from the monetary incentives that was provided at the end of the session. At the end of the experiment, all participants received the same amount of incentive bonus, which was $10.
Task.
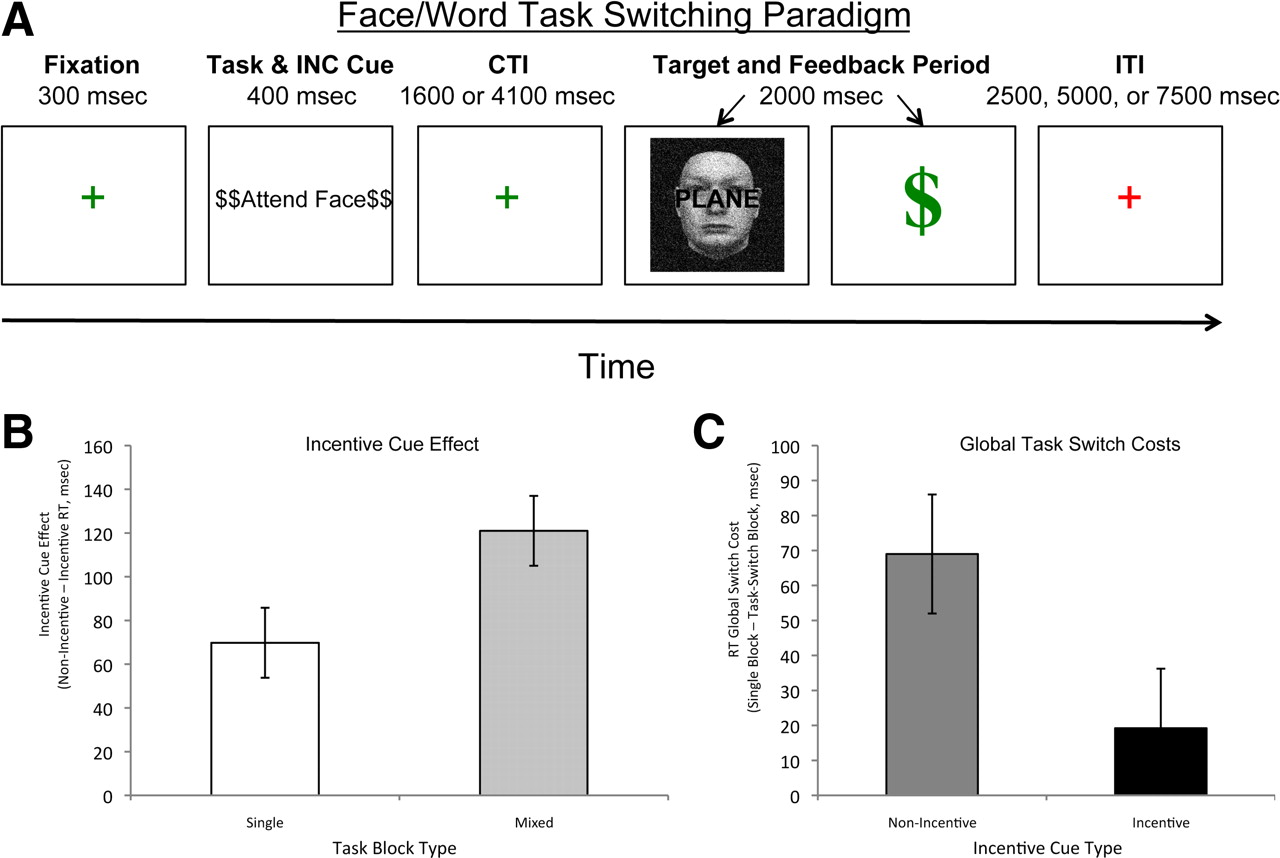
Participants engaged in a cued-task switching paradigm, structured as shown in Figure 1A. Two different classification tasks were performed on superimposed face–word stimuli: gender judgments (male or female) for the faces and syllable judgments (two syllable or not) for the words. The tasks and stimuli were adapted from those used in previous studies (Yeung et al., 2006; Savine et al., 2010). Stimulus combinations were created randomly from a bank of 144 faces (male and female), 72 two-syllable words, 36 one-syllable words, and 36 three-syllable words. The faces were stripped of hair and distorted with noise to equate perceptual difficulty between the words and faces.
All trials consisted of the following phases: fixation (300 ms), cue (400 ms), preparatory interval [also known as cue-to-target interval (CTI)] (variable duration; see below), target/response/feedback phase (2000 ms total), and intertrial interval (ITI) (variable duration; see below). During the cue phase, participants were informed of the task to be performed on the upcoming target (“Attend Face” or “Attend Word”). On incentive blocks, the cue also indicated the incentive value of the upcoming target, as described further below. The preparatory interval provided sufficient time (>1.5 s) to use the cue information to prepare for the target. In task-switching blocks, the two task cues were randomly intermixed, with the constraint that, on 50% of the trials, the cue was identical to the previous trial (task-repeat), and on the other 50%, the cue switched from the previous trial (task-switch). In single-task blocks, the same cue repeated on every trial in the block. Thus, on single-task blocks, the task cue provided no information (and thus was not critical for preparation).
During target presentation, participants were instructed to press one of two buttons (with the middle or index finger of right hand) to indicate classification of the task-relevant target dimension (either face or word), with response mappings for each task counterbalanced across participants. After the response, participants were presented with visual feedback indicating if an error was made (“Incorrect”) and, on incentive trials, whether reward was obtained (see below for details).
Procedure.
Participants first performed the task in a baseline behavioral session outside of the scanner before the scanning session. This session served to provide sufficient practice with the task and to provide a stable estimate of baseline performance. During this session, no incentives were provided, and participants had no knowledge regarding the future potential for incentives. These baseline blocks served to provide sufficient task practice for performance (and task learning) to reach an asymptote. Each block consisted of 48 trials, with the CTI fixed across trials at 1600 ms and the ITI fixed at 2500 ms.
In the fMRI scanning session of this study, participants performed 10 blocks of the task in separate scanning runs. The first two scanning runs consisted of single-task blocks, still performed in the absence of incentives (and with no knowledge regarding the future potential for incentives). On task-switching blocks, the task cue varied randomly from trial to trial, with each cue type (“Attend Face” or “Attend Word”) occurring with equal frequency. Single-task blocks were identical to task-switching blocks except that only a single task was cued throughout the entire block (and participants were informed of this before the start of the block). These scanning baseline blocks were included to set the reward criterion and were excluded from the primary data analysis described below. After this phase of the session, participants were informed that they would be performing additional experimental blocks but that, in these blocks, they would now have the potential to earn monetary bonuses on some trials based on their performance. This incentive phase consisted of two single-task blocks and six task-switching blocks, with each block consisting of 36 task trials for a total of 36 single-task non-incentive trials (SNG_NoInc), 36 single-task incentive trials (SNG_Inc), 108 task-switching non-incentive trials (SWT_NoInc), and 108 task-switching incentive trials (SWT_Inc). Condition order (single-task vs task-switching) was counterbalanced across participants. The trial structure during fMRI scanning was identical to that described above, except that CTI was 1800 ms on half the trials and 4300 ms on the other half, randomly intermixed. Additionally, the ITI was jittered between 2500 and 7500 ms in 2500 ms steps. This temporal jittering of the ITI allowed deconvolution of event-related fMRI responses. In addition to the 36 standard task trials, each block also included 12 partial (or cue-only) trials. In these trials, the target and feedback were not presented. Instead, the ITI began immediately at the end of the CTI (the start of the ITI was always indicated by a change in the fixation cross from green to red) (Fig. 1A). As described further below, the inclusion of these cue-only trials enabled fMRI analyses that isolated cue-related activation.
During the incentive phase, incentive value varied randomly on a trial-by-trial basis. Half of the trials were incentive trials, indicated by $$ symbols surrounding the task cue. The other half of the trials were non-incentive trials, indicated by XX symbols surrounding the task cue. Incentive cues varied orthogonally with task cues. Incentive cues were also present during the baseline blocks, but, in these blocks, participants were told that these symbols were irrelevant. This was done as a control to rule out that subsequent performance in the incentive blocks could be affected by novelty of the incentive cues or differences in low-level perceptual salience.
On incentive trials, participants had the potential to earn monetary rewards, based on task performance. Specifically, rewards were only obtained on trials in which the response was correct and faster than a prespecified reaction time cutoff, set individually for each participant based on their performance during the baseline scanning blocks. The cutoff was the 30th percentile (ordered from fastest to slowest) of correct response times (RTs) during these baseline trials. Because this cutoff was demanding and individually titrated, it ensured that each participant had to maintain optimal task performance to achieve high rates of reward during the incentive blocks. Moreover, the performance criteria was chosen to avoid ceiling effects such that, even with high motivation and performance, there would still be a substantial proportion of incentive trials that would not be rewarded.
Feedback was provided immediately after responses were made on incentive trials that indicated whether the reward was obtained. Specifically, when the incentive performance criteria were met, a large green dollar sign was presented after the response. The dollar sign did not correspond to a fixed amount of monetary reward, and participants were told only that when they received this feedback it indicated they had increased the amount of monetary bonus that they would obtain after the experiment. The ambiguity surrounding the exact meaning of the dollar sign was purposeful, to prevent participants from paying too close attention to the exact monetary earnings or for mentally updating and maintaining earnings in working memory. The use of a more abstract feedback cue, in terms of its relationship to monetary rewards, provided a stronger test of whether motivational incentives could modulate task-related brain activity, independent of other possible reward processing effects. Correct responses that did not meet incentive performance criteria (i.e., the RT cutoff) were followed by the words “Next Trial” as feedback. On non-incentive trials, regardless of reaction time, correct responses were followed by a change in color or size to target stimulus. In all conditions, incorrect responses were followed by the word “Incorrect” as feedback.
fMRI methods.
Functional images were acquired on a head-only Siemens 3 Tesla Allegra System. A pillow and tape were used to minimize head movement in the head coil. Headphones dampened scanner noise and enabled communication with participants. Both structural and functional images were acquired during the scanning session. Anatomical images were acquired using a magnetization-prepared rapid-acquisition gradient echo T1-weighted sequence. Functional images were acquired using an asymmetric spin-echo echo-planar sequence sensitive to blood oxygen level-dependent (BOLD) magnetic susceptibility (repetition time, 2.5 s; echo time, 25 ms; flip angle, 90°; slice thickness, 4 mm; in-plane resolution, 4 × 4 mm2; 32 slices). Functional images were acquired parallel to the anteroposterior commissure line allowing complete brain coverage at a high signal-to-noise ratio (Conturo et al., 1996). Each BOLD scanning run consisted of 228 scans (9.5 min duration) and started and ended with a fixation block (37.5 s), with the task performed through the rest of the block. The first four images in each scanning run were used to allow the scanner to ensure equilibrium of longitudinal magnetization and were discarded.
Visual stimuli were presented using PsyScope software (Cohen et al., 1993) running on an Apple PowerMac G4. Stimuli were projected to participants with a liquid crystal display projector onto a screen positioned at the head end of the bore. Participants viewed the screen through a mirror attached to the head coil. A fiber-optic, light-sensitive key press interfaced with the PsyScope Button Box was used to record participants' behavioral performance.
Data analysis.
Behavioral data were analyzed for incentive effects by conducting ANOVAs and t tests on error rates and RTs. All functional imaging data were preprocessed and statistically analyzed using in-house software (FIDL). Preprocessing involved temporal alignment of volume slices (to correct for asynchronous slice acquisition), normalization within each scanning run to a fixed image intensity value, and then correction for motion using a rigid-body rotation and translation algorithm (Friston et al., 1996; Snyder, 1996). Anatomical images were transformed into standardized atlas space (Talairach and Tournoux, 1988), using a 12-dimensional affine transformation (Woods et al., 1992, 1998). The functional data were then resampled into 3 mm cubic voxels, registered to the subject's anatomical images, and spatially smoothed with a 9 mm full-width half-maximal Gaussian kernel.
A general-linear model (GLM) approach (Friston et al., 1995) was used to estimate parameter values for event-related responses. Event-related effects were analyzed by estimating values for the various time points within the hemodynamic response epoch. The duration of this epoch was taken to be 22.5 s (11 scanning frames). This approach to GLM estimation (as opposed to a fit to predefined hemodynamic response function model) has been found to be critical in estimating complex trials or multi-event-related responses in rapid event-related designs. A first GLM coded the data at the trial level. In this model, each of the four trial types of interest was estimated with a separate set of regressors: (1) SWT_NoInc, (2) SWT_Inc, (3) SNG_NoInc), and (4) SNG_Inc. An additional set of regressors (of no-interest) coded for cue-only and error trials. A second GLM focused only on the task-switching condition but used the cue-only trials to independently estimate cue- and target-related activity, separately for incentive and non-incentive trials (Ollinger et al., 2001). As described more fully below, this model also used an additional set of regressors that enabled examination of the modulatory effects of behavioral performance (fast vs slow responses) on brain activity and to control for performance differences between incentive and non-incentive trials. The data from these GLMs were analyzed using a three-stage approach, which enabled examination of task-switching and incentive effects in progressively greater detail but controlled for circularity or non-independence biases in the examination of regions of interest (ROIs) (Kriegeskorte et al., 2009).
In the first stage of analysis, we identified voxel clusters sensitive to task-switching and incentive effects within two discrete networks of theoretical interest: the CCN and the reward network (REW). The CCN ROI mask consisted primarily of frontal and parietal regions, and its coordinates were defined from the coordinates of regions derived from two published meta-analyses (Wager and Smith, 2003; Owen et al., 2005). We have used this identical spherical ROI mask successfully in previously published studies (Emery et al., 2008; Fales et al., 2008). The REW defined the core set of brain regions associated with reward and affect, primarily comprising subcortical regions such as the ventral and dorsal striatum and amygdala but also including cortical structures such as the posterior insula and ventral orbitofrontal cortex (Knutson et al., 2003). REW regions were hand drawn according to anatomical landmarks, as well as coordinates provided in previously published studies of reward effects (Knutson et al., 2001, 2003; O'Doherty et al., 2003, 2004; Kringelbach and Rolls, 2004; Nitschke et al., 2006; Ahsan et al., 2007; Jensen et al., 2007; Kable and Glimcher, 2007; van den Bos et al., 2007). We have successfully used this mask in a previously published study as well (Beck et al., 2010). The exact coordinates for both CCN and REW can be found in supplemental Tables S1 and S2 (available at www.jneurosci.org as supplemental material).
These networks were then used as masks to constrain analysis to only those voxels that were theoretically expected to be most strongly associated with cognitive control and reward processing. We identified voxel clusters from within these masks that showed particular cognitive control and incentive effects of interest. To identify these patterns, we constructed multiple contrasts, and a voxel cluster was only identified if it simultaneously satisfied all contrasts. Moreover, a statistical significance threshold of z = 3 and a minimum cluster size of 19 voxels was chosen to protect against false positives, even within the smaller networks of interest. These contrasts each involved the GLM estimates for different trial conditions, averaged across time points 2–6, to capture the primary processing period of the trial from the time the incentive/task cue is presented to the time of target processing (after accounting for the ∼3–6 s lag in the hemodynamic response). A first set of contrasts identified voxel clusters sensitive to pure task-switching demands, i.e., under non-incentive conditions: SWT_NoInc > SNG_NoInc and SWT_NoInc > Fixation. A second set of contrasts identified voxel clusters sensitive to pure incentive effects, i.e., under non-switching conditions: SNG_Inc > SNG_NoInc and SNG_Inc > Fixation. All subsequent analyses were conducted at the ROI level exclusively on regions identified in this first stage of analysis. However, to examine whether any additional regions outside of the CCN and REW showed similar effects, a supplementary whole-brain exploratory analysis was also conducted. The same set of contrasts were used, but each contrast was first false discovery rate (FDR) corrected at p = 0.05. This more stringent statistical threshold was used to guard against false-positive identification associated with multiple comparisons in whole-brain analysis. No additional regions were identified outside of the CCN or REW network that met this criterion.
In the second stage of analysis, ROIs identified in the first stage were interrogated to examine the time course and combined effects of incentives and task-switching. Each ROI was subjected to a repeated-measures ANOVA that included the factors of incentive (Inc vs NoInc), task-switching (SWT vs SNG), and time (time points 2–6). Specifically, this analysis tested whether ROIs identified based on pure task-switching effects would also be modulated by incentives and, conversely, whether ROIs identified based on pure incentive effects would also be sensitive to task-switching. Furthermore, by including time as an additional factor, it was possible to tell whether incentive and task-switching modulation had different temporal dynamics. An additional analysis conducted in this second stage tested for between-subjects correlations between activation and task performance.
The third stage of analysis further examined the dynamics of task-switching and incentive effects through the use of the second GLM model, which separately estimated cue versus target-related activity in incentive and non-incentive trials. Because this GLM was lower powered and only estimated for task-switching trials, it was only applied at the ROI level and only to the ROIs that had been found to show effects of incentive in either the first or second stage. A key aspect of the analysis was to determine more rigorously whether incentive effects during task-switching occurred during the cue or target period. A second key aspect of the analysis was to examine and control for behavioral performance differences between incentive and non-incentive trials. This was done via another set of regressors that coded for the modulatory effect of behavioral performance on event-related activation dynamics. The performance regressor was coded as categorical variable on each full trial that indicated whether the response was faster (dummy coded as 1) or slower (dummy coded as 0) than the reaction-time cutoff that specified whether reward was obtained (and which was also individually set for each participant based on baseline performance; see above). Error trials were again coded separately with regressors of no interest, so this analysis only applied to correct trials. Importantly, the same cutoff was applied to non-incentive as well as incentive trials, to match the two trial types on behavioral performance effects so that these would be controlled for when testing for effects of incentives.
Results
Behavioral results
The CTI factor in this experiment was not found to be significant in any analyses (including interactions). Consequently, for the results reported below, all of the analyses have been collapsed across this factor.
We first observed that, even with a demanding reward criterion, performance was enhanced on incentive trials. Specifically, the average reward rate was 65% (range, 52–84%), which was much higher than the predicted value of 30% reward rate, based on baseline performance. Nevertheless, as predicted, the reward rate was significantly below ceiling, such that analyses comparing rewarded with nonrewarded incentive trials could also be conducted.
Additional, more direct enhancement effects of incentives were also observed for both errors and RT. On incentive relative to non-incentive trials, accuracy was 2.7% higher (F(1,15) = 4.58, p < 0.05) (Table 1) and RTs were 94 ms faster (F(1,15) = 26.15, p < 0.001). It is important to note that these effects were specific to the incentive condition, although the same incentive cues were present during the baseline block (when participants were naive to their meaning). Specifically, during the baseline blocks, performance was statistically identical on trials with incentive cues versus non-incentive cues, indicating that such cues only have effects on performance when they are motivationally meaningful (see section below).
Task-switching behavioral performance
Additional analyses examined the relationship between incentive and task-switching. Error rates did not interact with task-switching (F(1,15) = 1.6, NS). However, for RTs, a significant incentive × task-switching interaction was observed (F(1,15) = 11.38, p < 0.001). Specifically, greater incentive cue facilitation was observed in task-switching blocks (121 ms) relative to single task blocks (70 ms) (Fig. 1B), indicating a selectively stronger incentive benefit when cognitive control demands were highest. Additionally, when expressing the data in terms of switching costs (task-switching − single-task), it was found that, although these costs were reliable on non-incentive trials (69 ms, t(15) = 2.20, p = 0.05), they were no longer significant on incentive trials (19 ms, t(15) = 0.75, NS) (Fig. 1C). This reduction in switch costs replicates our previous behavioral data (Savine et al., 2010) and provides evidence that incentive motivation enhanced the ability to use cognitive control and flexibly switch task sets.

A, Paradigm description for task-switching. On each trial, a fixation cross appeared on the screen (300 ms), followed by a task cue (400 ms), a CTI screen with a (green) fixation cross (1600 or 4100 ms), and then presentation of the target stimulus and feedback for the response (2000 ms total). The end of the trial and start of the ITI (2500, 5000, or 7500 ms, jittered) was indicated by the fixation cross changing color (red). B, Incentive cue analysis. On incentive cued trials, performance improved in single-task and mixed-task conditions, with the largest incentive benefits occurring in the mixed-task condition. Thus, incentive cue effects were largest when cognitive control demands were highest. C, Switch cost analysis. Significant switch costs were present on non-incentive trials but not on incentive trials. This attenuation of switch costs suggests that incentive motivation enhances cognitive control, specifically the flexible switching of task set or goal information.
A final behavioral analysis examined whether the incentive facilitation effects were dependent on the reward outcome on the previous incentive trial. When incentive trials were divided into those that were associated with a rewarded versus non-rewarded outcome (e.g., performance was below the reward cutoff), we calculated the conditional probability of obtaining the reward on trial n, if the previous trial n − 1 was rewarded. This probability was 62%, which is not significantly different from the rewarded rate of 65% on incentive trials overall. Thus, incentive trial outcomes seem to have a negligible effect on task performance of subsequent incentive trials (F < 1). In other words, the observed facilitation effect appeared to be related to the potential for obtaining reward, rather than dependent on success or failure in obtaining reward on a previous incentive trial. This suggests that analyses of cue-related incentive effects are unlikely to be contaminated by the residual processes associated with the feedback from preceding trials.
Preexposure of the incentive and non-incentive cues
In a baseline session outside of the scanner, as well in the first two baseline blocks in the scanner session, task-switching was performed in the absence of incentives. Importantly, in these naive blocks, the task cue was always flanked by what would be given value as non-incentive or incentive cues in the future blocks XX or $$. This was done to ensure that pure perceptual novelty would not be able to account for the incentive effects observed. It is possible, however, that the differences in trial type had effects even before the incentive instructions were given. To test this possibility, a 2 × 2 ANOVA with factors of task-switching (SWT vs SNG) and flanking cue ($$ vs XX) was conducted for error rates and response time to test for these potential effects within the baseline blocks. Neither error rates nor response times demonstrated main effects or interactions of flanking cues in the baseline blocks (F < 1), providing evidence that the cues did not bias performance when they were not ascribed an incentive value. Furthermore, there did not appear to be latent inhibition (an initial suppressing of incentive cued benefits) affecting performance benefits during the incentive blocks. There was instead an incentive-based benefit in RT performance even on the very first 10 incentive $$ cued trials of the first incentive block relative to the last 10 non-incentive $$ incentive cued trials of the last baseline block (924–708 ms), and the incentive cue effect did not vary significantly in magnitude for RT from the first to the last incentive block (F < 1), with error rates staying constant as well (F < 1). Thus, we can be confident that behavioral performance improvements observed on trials with high motivational value were attributable to enhanced cognitive control.
Imaging results
First-stage analysis: identification of pure task-switching and incentive effects
We first identified regions that showed increased activation associated with pure task-switching effects (i.e., using only non-incentive trials). Within the CCN, 10 regions were identified, including the bilateral DLPFC, bilateral inferior parietal lobule (IPL), dorsal anterior cingulate/presupplementary motor area (dACC/pre-SMA) as well as anterior and posterior PFC regions, such as the inferior frontal junction (IFJ) (Table 2; Fig. 2, red). This network of regions strongly replicates previous neuroimaging results of task-switching (Braver et al., 2003; Badre and Wagner, 2006; Crone et al., 2006; Ruge and Braver, 2007; Ruge et al., 2009). In contrast, within the REW, no regions were identified that showed significant effects of pure task-switching effects at the statistical thresholds used.
Summary of significant regions and effects

Regions of interest. The regions in red indicate the components of the CCN that showed significantly increased activation associated with task-switching. The regions in green indicate the components of the REW that showed significantly increased activation associated with reward incentives.
A complementary analysis was then conducted to identify regions that showed increased activation associated with pure incentive effects (i.e., under non-switching conditions). Within the CCN, no regions were found that showed such effects. In contrast, within the REW, three regions were identified: the bilateral dopaminergic midbrain, and a ventromedial PFC region often described as paracingulate cortex, because it is in the transitional zone between the rostral anterior cingulate and medial frontal pole (hereafter termed vmPFC/pACC; (Amodio and Frith, 2006; van den Bos et al., 2007). Although incentive effects were not observed in the ventral striatum or nucleus accumbens at the primary statistical threshold, such effects were found when this threshold was lowered to z = 2.4. We report this finding because of the typical observation of ventral striatal activity associated with reward processing (McClure et al., 2004; Montague et al., 2006; Haber and Knutson, 2010), but because of the questionable significance of the effect, do not include this region in additional analyses.
There may be a few reasons why incentive effects in the ventral striatum may not have activated regions in the REW as robustly as in other reward paradigms. The reward in this study was more abstract, because participants were not explicitly provided a translation between reward feedback on each trial and a specific monetary amount obtained. Furthermore, the incentive cue did not directly indicate future reward, because reward attainment is contingent on specific performance criteria, which in turn are contingent on successful high-level cognitive processing. The indirect relationship between incentive cues may have weakened the relationship between these cues and activation within reward regions. Additionally, the incentive cues were presented simultaneously with task cues, which may have further weakened their “pure” associative relationship with reward, yet it is important to point out that robust incentive-related activation of the bilateral dopaminergic midbrain and vmPFC/pACC and a trend toward activation in the ventral striatum were observed. Thus, we can be confident that task paradigm is sufficiently sensitive to detect incentive effects on brain function.
Together, these results suggest the potential anatomical dissociability of effects related to cognitive control and reward processing, because pure task-switching effects were selectively associated with activation in the CCN, whereas pure incentive effects were selectively associated with activation in the REW.
Second-stage analysis: combined effects of incentives and task-switching on activation dynamics
The next analysis was conducted at the ROI level on regions identified in the first stage. The time courses of each of the 13 identified ROIs were extracted and subjected to repeated-measures ANOVAs, including the factors of incentive (Inc vs NoInc), task-switching (SWT vs SNG) and time (time points 2–6). These ANOVAs were conducted to test whether activation in the identified ROIs were sensitive to the combined influence of incentives and task-switching. It is important to note that this second-stage analysis is not independent of the first stage, because it uses some of the same conditions. However, it is negatively rather than positively biased, according to the criteria discussed by Kriegeskorte et al. (2009). This means that pure task-switching ROIs identified in the first stage will be biased to have large task-switching effects on non-incentive trials and thus, by regression to the mean, should show reduced task-switching effects on incentive trials (the so-called “overfitting” effect described by Kriegeskorte et al., 2009). Put another way, the “deck is stacked” against detecting that task-switching activation is greater under incentive than non-incentive conditions. Thus, detection of such effects would constitute a more rigorous confirmation of the hypothesis.
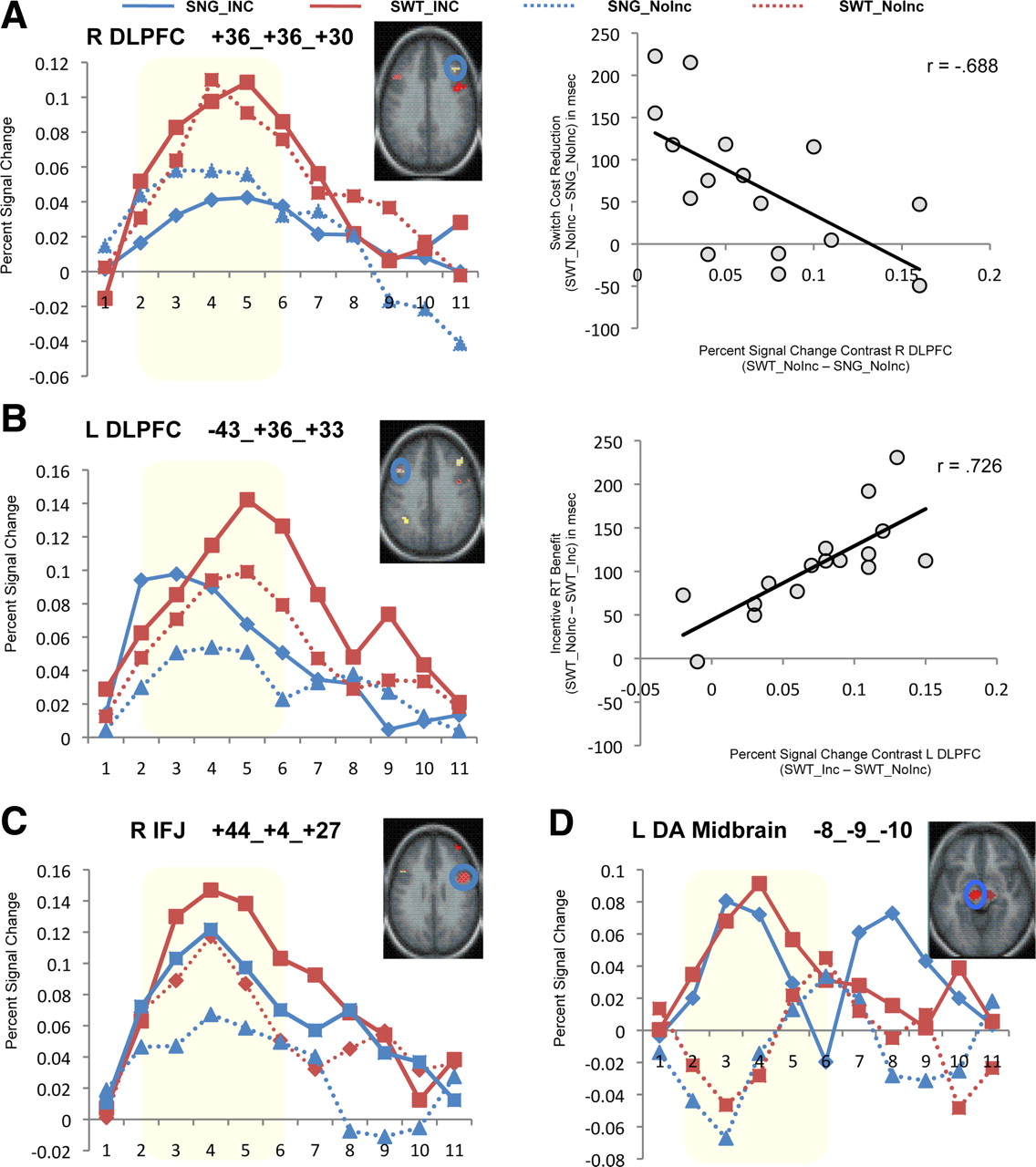
In 4 of the 10 CCN regions identified as showing task-switching effects, there were no additional incentive effects observed, such that only the main effect of task-switching was significant. These regions included the right DLPFC (Fig. 3A, left), occipital cortex, and cerebellum. In a fifth region, the right anterior PFC, incentive effects were inconsistent and only present on single-task conditions (i.e., not during task-switching). Critically, in another four regions, including the left DLPFC, dACC/pre-SMA, and bilateral IPL, significant incentive-related increases in activation were observed, in addition to the switching-related increases. Thus, switching-related activation in these regions was increased on incentive trials. Incentive effects were also observed on single-task trials; however, the nature and time course of these incentive effects were different in the two conditions (switching vs single-task), resulting in incentive × task-switching × time interactions. For example, in the left DLPFC (F(4,12) = 4.57, p < 0.05) (Fig. 3B, left), incentive induced increases in activation occurred only early in the trial (during incentive cue period) in the single-task block but persisted in task-switching trials, suggesting that incentives had a prolonged effect under task conditions in which enhancement of cognitive control is required for optimal performance.

Full-trial analyses illustrating time course of activation during single-task and task-switching trials under incentive versus non-incentive conditions. A, Right DLPFC. This CCN region showed a main effect of task-switching but not incentives (left). Increased task-switching-related activity (SWT–SNG) in this region is correlated with reduced RT switch costs (right). B, Left DLPFC. Representative CCN region showing a task-switching × incentive × time interaction. Incentive effects on single-task trials occur only early in the trial (during incentive cue period) but persist in the mixed-block trials, suggesting a modulatory effect on control processes (left). Increased activity in this region on incentive trials during task-switching (SWT_Inc − SWT_NoInc) correlated with greater incentive-related RT facilitation (right). C, Right IFJ. This CCN region exhibited main effects of both task-switching and incentives but no interaction. D, Left dopaminergic midbrain. Representative REW region showing a main effect of incentives but no effects of task-switching. In all regions, highlighted window indicates time points (2–6) included in statistical analysis, corresponding to the primary processing period of the trial from incentive/task cue to target response, after accounting for hemodynamic lag. L, Left; R, right.
In a final region, the right IFJ, there was also a significant incentive effect (F(1,15) = 4.91, p < 0.05), but the nature and dynamics of the effect was the same on single-task and task-switch trials, indicating an additive pattern (Fig. 3C). This finding, of IFJ sensitivity to both task-switching and incentive effects, extends previous work demonstrating that the IFJ is involved in the activation of task representations (Brass et al., 2005). The additive pattern of incentive and task-switching effects suggest that these two variables are treated as equivalent signals in the IFJ and may indicate that this brain region may be important for updating task representations based on cues that indicate high salience or priority of this information (e.g., either switch or incentive cues).
In contrast to the incentive effects observed within task-switching-sensitive regions in the CCN, ANOVAs on the incentive-sensitive regions in the REW did not show any additional effects of task-switching. Thus, in all three regions, only the incentive effects were significant. The left dopaminergic midbrain is shown as a representative example demonstrating the main effect of incentive (Fig. 3D) (F(4,12) = 7.79, p < 0.01) but a complete absence of any task-switching effect. These effects are consistent with the hypothesis that REW regions are primarily involved in coding reward-related processes on incentive trials rather than in integrating such information with cognitive control demands. Nevertheless, it is important to qualify that the null effect of task-switching in incentive-related REW regions should be treated with caution, because it could be attributable to the lower power to detect such effects associated with the negative bias introduced in the selection criteria, as described above.
Hemispheric specialization in DLPFC
The contrasting patterns of incentive effects observed in the CCN regions associated with task-switching suggest differential functional roles in task behavioral performance. In particular, the right DLPFC showed sensitivity to task-switching but not incentive effects, whereas the left DLPFC was sensitive to both factors. To test whether these patterns in left and right DLPFC were truly dissociable, we conducted a direct statistical comparison of the two regions in a 2 × 2 × 2 ANOVA, with region, incentive, and task-switching as factors. The three-way interaction was significant (F(1,15) = 4.73, p < 0.05), indicating that the incentive × task-switching interaction found in the left DLPFC region was statistically different from the effect observed in the right DLPFC region.
We further examined the differential functional patterns in right and left DLPFC by examining the between-subjects correlation between activation and task performance. In the right DLPFC, smaller behavioral switch costs (SWT–SNG) were associated with larger switch-related activation increases (i.e., SWT_NoInc − SNG_NoInc % BOLD signal change), and this was true on both non-incentive (NoInc, r(14) = −0.688, R2 = 0.47, p = 0.005) (Fig. 3A, right) and incentive (Inc, r(14) = −0.549, R2 = 0.30, p < 0.05) trials. However, incentive-related performance benefits (NoInc − Inc) were not associated with corresponding incentive-related activation effects in either task-switching (SWT_Inc − SWT_NoInc % BOLD signal change; r(14) = −0.128, R2 = 0.02, NS) or single-task (SNG_Inc − SNG_NoInc % BOLD signal change; r(14) = −0.106, R2 = 0.01, NS) conditions. In contrast, for the left DLPFC region, incentive-related performance-activation relationships were present. A larger incentive-related facilitation of performance in the task-switching condition (i.e., SWT_NoInc − SWT_Inc RT; reverse coded so that facilitation effects are in the positive direction) was associated with a greater incentive-related activation increase in the same conditions (i.e., SWT_Inc − SWT_NoInc % BOLD signal change; r(14) = 0.726, R2 = 0.53, p = 0.001) (Fig. 3B, right). This incentive effect was not observed in the single-task condition (r(14) = 0.263, R2 = 0.07, NS), suggesting that it was selectively present when the high cognitive control demands of task-switching were involved. Moreover, in the left DLPFC, like the right DLPFC, there was also a relationship between switch-related activation and behavioral switch costs that was present even under non-incentive trials (SWT_NoInc − SNG_NoInc % BOLD signal change; r(14) = −0.637, R2 = 0.41, p < 0.01).
Together, these distinct activation and correlation patterns provide convergent evidence that the left and right DLPFC make different functional contributions to task performance under motivated task-switching conditions.
Third-stage analysis: isolating incentive effects on cue versus target-related activity during task-switching
The previous analyses identified a set of regions for which activation increased on incentive relative to non-incentive trials. Because this analysis was conducted at the whole-trial level, it did not enable discrimination of whether such incentive effects were associated with the cue period, target/feedback period, or both. To address this question, a second GLM was used, which enabled independent estimation of cue versus target-related activity separately for incentive and non-incentive trials but focusing selectively on the task-switching condition (for full a description, see Materials and Methods). The GLM estimates of these different effects (Cue/Target × Inc/NoInc) were extracted at the ROI level for each of the identified regions that showed incentive-related enhancements in the second-stage analysis (five CCN and three REW ROIs). It is important to note that this third-stage analysis is unbiased by the results of the previous stages, because it concentrates exclusively on the task-switching condition and only tests whether the previously established incentive effects can be isolated to either cue or target periods.
In all of these eight regions, incentive-related activation increases were associated with the target event (Table 2; Fig. 4A). In addition, there was also significant incentive-related activation at the time of the cue for the right IFJ (F(1,15) = 6.86, p < 0.05) and left DLPFC (Fig. 4B) (F(1,15) = 7.08, p < 0.05) regions. This suggests that, in these two regions, the incentive information provided at the time of the cue may have been encoded and used to enhance task-related preparation as a means of maximizing rewards.

A, Activity during target/feedback period. The majority of regions identified [including CCN regions such as left (L) DLPFC and REW regions such as left dopaminergic midbrain shown here] exhibited greater activity at the time of the target on incentive trials. B, Cue-related neural activity. The left DLPFC (left) exhibited greater cue-related activation in response to the processing of incentive cues compared with non-incentive cues. Regions in the REW such as the left dopaminergic midbrain (right) do not show such differences in cue-related activity. C, Trial-outcome (reward) effects during target/feedback period. Two REW regions, including the left dopaminergic midbrain (right), demonstrated increased activation specifically on rewarded incentive trials, consistent with dopaminergic effects in response to achieving performance contingent, and thus unpredictable, rewards. D, Behaviorally linked cue activity. The left DLPFC (left) exhibited greater cue-related neural activity on trials that subsequently led a rewarded outcome as a result of optimal performance. No such effects occurred on matched performance trials that were not rewarded, indicating a specific effect of the incentive cue.
An important residual issue relates to the relationship between incentive cues, behavioral performance, and trial outcomes. In particular, behavioral performance on incentive trials was significantly better than that on non-incentive trials (in terms of both accuracy and reaction time). Furthermore, only on incentive trials is optimal trial performance (in terms of an accurate and fast response) associated with a meaningful outcome (i.e., whether reward is received or not). This suggests that increased cue- and target-related activation on incentive trials could either be related to the greater motivational salience of these trials (i.e., related to the performance-contingent reward expectancy signaled by the incentive cue) or to the improved behavioral performance, independent of any motivational salience effects. To tease apart these issues, the second GLM model included a separate set of behavioral regressors that categorically coded for trial performance. This categorical coding defined trials as either being faster or slower than the reaction time cutoff used to determine whether the trial would be rewarded or not (only correct trials were included in the analysis, because error trials were coded with separate regressors of no interest). Behavioral performance coding was applied not only to incentive trials but also identically to non-incentive trials as well. As a consequence, the behavioral performance regressors provided a means of examining behavioral performance effects (optimal vs poor performance or fast vs slow reaction time) in a matched manner across incentive and non-incentive trials. However, only in incentive trials is this distinction motivationally meaningful to the participant, because on incentive trials it defines whether the trial will be rewarded or not (whereas on non-incentive trials the distinction is arbitrary and irrelevant to participants). As such, any selective effects of incentive related to behavioral performance can be interpreted more strongly as being related to trial outcome (rewarded or not) and/or motivational salience (high or low).
We first examined the behavioral performance/trial outcome effect on target-related activity (Fig. 4C). Because reward feedback was provided close in time after the target presentation (and the associated response), activation related to outcome occurring to target events can be reasonably interpreted to include the effects of such feedback. We found that activation in both dopaminergic midbrain regions was increased around the time of the target selectively on rewarded trials but not on matched non-incentive trials with similar behavior performance profiles [left dopaminergic midbrain: F(1,15) = 7.06, p < 0.05 (Fig. 4C, right); right dopaminergic midbrain: F(1,15) = 5.72, p < 0.05]. The fact that activity in these regions was modulated around the time of reward feedback on incentive trials is consistent with the extensive literature suggesting dopaminergic neural activation associated with unpredictable rewards (Mirenowicz and Schultz, 1994; Schultz, 1998; Berns et al., 2001).
The analysis of cue-related activation also found unique patterns of activation associated with subsequent trial outcomes. Specifically, in the left DLPFC region, increased cue-related activation was found on incentive trials associated with optimal behavioral performance/rewarded outcome (Fig. 4D, left) (t(15) = 3.02, p < 0.01). This effect was selective to incentive trials, because there was no effect of behavioral performance on cue-related activation during non-incentive trials. Finally, behavioral performance/trial outcome was not did not modulate target-related activity on either incentive or non-incentive trials in this region (Fig. 4C, left) (t(15) = 0.483, NS), and a similar absence of target-related modulation was found in the other PFC regions as well.
These results suggest that, in the left DLPFC, the response to task cues is selectively increased when the cue indicates that the trial is of high motivational salience. Moreover, the behavioral performance effect indicates that the cue-related activation increase cannot be attributed to performance differences between incentive trials but, conversely, that the magnitude of cue-related activity on incentive trials is predictive of whether the trial will be subsequently rewarded. This trial-by-trial relationship is consistent with the hypothesis that the presence of incentive cues may have facilitated successful encoding and preparatory maintenance of task-related information needed to enhance cognitive control. As a final convergent test of this hypothesis, we examined the relationship between the reward-related incentive cue response and between-subjects variation in the two primary behavioral measures indexing incentive facilitation of performance: reward rate and the incentive cue effect. A significant positive correlation was found with reward rate (r(14) = 0.574, R2 = 0.33, p < 0.05), and a similar trend-level correlation was found with the behavioral incentive cue effect (r(14) = 0.436, R2 = 0.19, p = 0.08).
Discussion
The results of this study support two main conclusions. First, they provide new evidence of a relatively selective motivational influence on brain activity related to cognitive control. Only a subset of regions within the neural cognitive control network demonstrated sensitivity to both task-switching and incentive information. Conversely, regions showing pure incentive effects (i.e., under low cognitive control demands) were in the REW network rather than CCN, and were not sensitive to task-switching. This pattern of results suggest the interaction of motivation and cognition occurs within a fairly narrow set of specialized brain regions rather than as a more global, arousal-type process. Second, the study provides strong evidence that the behavioral performance improvements observed on trials with high motivational value were attributable to enhanced cognitive control. Specifically, increased switch-related activation in DLPFC was associated with lower behavioral switch costs. Moreover, incentive enhancement of task-switching activation was associated with the enhanced behavioral performance observed on incentive trials. Critically, the effects were preparatory, because selective activation increases during task cue presentation occurred on incentive trials and predicted both trial-by-trial reward attainment and between-subjects differences in average reward rate. These results suggest that incentive signals facilitated the encoding and utilization of task cue information, thus optimizing processing and preparation before future targets.
The current study replicates previous results demonstrating increased task-switching-associated activation in regions of the brain cognitive control network, including DLPFC, IFJ, dACC/pre-SMA, and IPL (Sohn et al., 2000; Brass et al., 2005; Crone et al., 2006; Ruge and Braver, 2007; Sakai, 2008). Moreover, we extend these results by demonstrating that motivational incentives further increase activation in these regions and enhance behavioral performance. For example, new evidence was provided that the IFJ may activate or update task-set information on the basis of either task or incentive cues. The activation pattern suggests that this region may treat both types of cues as equivalent signals indicating the high salience or motivational priority associated available task-set information. Under standard switching conditions, this is because task cues provide critical information about the task set that is relevant for the current trial, whereas under single-task conditions, incentive cues also provide information about the importance of the current task set, which may enable a refresh or strengthening of the representation. Additionally, the findings indicate that DLPFC regions exhibit the strongest associations between increased activation and enhanced performance. Although not negating the potentially critical control functions played by the IPL and dACC/pre-SMA during task-switching (Luks et al., 2002; Brass and von Cramon, 2004; Wager et al., 2004; Bunge et al., 2005; Crone et al., 2006), the current findings indicate that, at least under salient motivational contexts, between-subjects variation in task performance is most strongly predicted by between-subjects variation in DLPFC activation patterns.
The left dorsolateral PFC in particular appears to serve as a key neural locus for the integration of information indicating motivational salience and cognitive control demand. This result is highly consistent with previous monkey neurophysiological studies showing such effects in single PFC neurons (Leon and Shadlen, 1999; J. Watanabe et al., 2002; M. Watanabe et al., 2002) and particularly the Leon and Shadlen (1999) finding that reward cues sharpened PFC representations in working memory. Interestingly, the effects of cognitive control demand and motivation were hemispherically dissociated within DLPFC in the current study. Although activation related to task-switching and behavioral switch costs was found in PFC bilaterally, only left DLPFC was sensitive to motivational incentives or associated with incentive performance modulations. This is consistent with work showing a left-hemisphere bias in lateral PFC activity during task-switching (Brass and von Cramon, 2004; Badre and Wagner, 2006). Lateralization effects may also depend on whether transient (i.e., event-related) or sustained activation is examined (Braver et al., 2003; Jimura and Braver, 2010; Jimura et al., 2010).
A striking finding was that task performance predictions could be based not only on the general trial-related response in left DLPFC but selectively from cue-related activation on incentive trials. This is a very specific result, because cue-related enhancement was not observed on non-incentive trials associated with successful behavioral outcomes (fast and accurate performance) but not rewards. Furthermore, greater cue-related activity in this region predicted both within-subject (trial-by-trial) and between-subjects variation in performance, because individuals showing greater cue-related left DLPFC activity also demonstrated the largest incentive-related performance enhancements. Thus, preparatory activation in left DLPFC may be the single best neural predictor of successful, motivated cognitive control. This result supports the hypothesis that cue-related left DLPFC activity indexes task set or goal strength and that motivational priority or salience significantly modulates this activation strength during task performance.
These findings have important implications when considered in relation to the dual mechanisms of control (DMC) framework (Braver et al., 2007). The DMC framework suggests that cognitive control consists of two modes: proactive (anticipatory and sustained control during cue encoding and maintenance periods) and reactive (control engaged just-in-time during task probe or response periods). Differential neural activity dynamics dissociates these modes, but they may flexibly shift within the same neural region (Braver et al., 2009). Thus, the results suggest that motivational incentives must be associated with a shift toward proactive cognitive control (increased cue-related activation) to obtain optimal performance benefits. Similar conclusions have been drawn from examining the effects of motivational incentives in other cognitive control and attentional tasks (Engelmann et al., 2009; Jimura et al., 2010). Indeed, performance enhancement was associated with facilitation of cue-related, but not target-related, processing in dorsolateral PFC. An additional test of the DMC framework would be to present incentive cues at target rather than task cue presentation, because the framework would predict that such incentive signals could only impact reactive and not proactive control. Thus, increased target-related activation would be expected in the same regions of the brain cognitive control network but without the associated enhancement in task-switching performance.
We also observed pure reward incentive effects (insensitive to task-switching demands) within the dopaminergic midbrain and ventromedial PFC (vmPFC/pACC). These regions are posited as critical components of brain reward circuitry, because current models suggest that they track subjective reward value, and are important for encoding reward values associated with possible future outcomes, especially when contextualized by higher-order expectancies and cognitive processes (Hare et al., 2008, 2009). Furthermore, extensive work has demonstrated that the midbrain dopaminergic system exhibits phasic responsivity to both unpredicted rewards and reward-predicting cues (Schultz et al., 1997; Schultz, 2000).
A question remains as to whether the robust responses to reward incentives found in these brain regions are the source of the motivational effects on task-switching performance. The midbrain dopaminergic system has been implicated in such interactions. First, the DA system appears to support motivational as well as reward-prediction functions (Berridge, 2007), because phasic dopaminergic reward cues predict performance enhancements (Schultz, 2002; Satoh et al., 2003). Second, DA release occurs during cognitive tasks (Watanabe et al., 1997), with phasic activation occurring to task cues indicating reward potential (Ljungberg et al., 1992). Third, dopamine (DA) modulates preparatory PFC activity during active maintenance periods in working memory and other cognitive control tasks (Sawaguchi and Goldman-Rakic, 1991; Williams and Goldman-Rakic, 1995; Cools, 2008).
Braver and Cohen (1999, 2000) developed a specific computational model of dopamine incorporating these findings that may explain the observed motivational influences on cognitive control. The model specifies that phasic DA activity in response to reward-predicting task cues enables the appropriate updating of goal representations in PFC while allowing for stable maintenance of such representations when phasic DA responses are absent. Applied to the current results, the model suggests an association between incentive cues and increased phasic dopamine release within the PFC, enabling optimal and efficient updating of task-set information during task-switching (Müller et al., 2007). Nevertheless, some aspects of the observed results are inconsistent with this account, because target stimuli and rewards, but not cues, were associated with increased dopaminergic midbrain activity. One possible explanation is that our design did not enable separate estimation of incentive cue versus task cue activation, perhaps weakening the ability to detect incentive cue effects on dopamine activation.
Alternatively, recent theoretical models have posited that tonic DA may mediate changes in motivational salience and subsequent incentive effects (Niv et al., 2007). Tonic DA would then serve as a signal of a reduced “opportunity cost” for effortful behaviors (Salamone et al., 2007), leading to overall increases in vigor (or speed) of response or action selection. Within the current experimental context, tonic DA may increase during performance of blocks with high incentive value, providing a baseline shift that enhances the ability for cue-related encoding and maintenance of task-set information in working memory. Such an effect would be consistent with many neurocomputational models of DA effects in PFC, in which tonic DA stimulation (primarily through D1 receptors) increases the robustness and stability of active maintenance processes (Durstewitz et al., 2000; Cohen et al., 2002).
The distinct accounts regarding the functional roles of tonic and phasic DA suggest that isolating sustained and transient modulatory effects of reward incentives on cognitive control may be desirable. Indeed, in previous behavioral and neuroimaging studies with other cognitive control tasks, both trial-related (transient) and contextual (tonic, state-related) motivational components have been observed (Locke and Braver, 2008; Beck et al., 2010; Jimura et al., 2010; Savine et al., 2010) using appropriate experimental designs. Thus, applying these approaches (such as mixed transient/sustained fMRI methods (Visscher et al., 2003) with the current task context may adjudicate whether reward incentive effects during task-switching are linked to differential phasic versus tonic activation in the dopaminergic midbrain.
Conclusions
In summary, our findings not only indicate that motivational incentives enhance task-switching performance but also that increased activation in left DLPFC may signal the integration of incentive and task information needed to achieve optimal task performance. Incentive-related effects in left DLPFC were most prominent and predictive of task performance during cue-related processing, suggesting that the motivational benefits were preparatory in nature. Thus, the results suggest that the maximal benefits of incentive information occur through a proactive cognitive control process that enhances the encoding and maintenance of task set information. The engagement of midbrain dopaminergic regions on incentive trials, and during periods of reward feedback, is consistent with a role for the dopaminergic system as an energizing source of the motivation–cognitive control interaction. Together, the results highlight the utility of motivational manipulations in elucidating the neural mechanisms of cognitive control.
Footnotes
This work has been supported by National Institutes of Health R01 Grant 2R01MH066078 (T.S.B.).
- Correspondence should be addressed to Adam C. Savine, Department of Psychology, Campus Box 1125, Washington University in St. Louis, St. Louis, MO 63139. acsavine{at}artsci.wustl.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}