
One of the main characteristics of neurons is their highly structural and functional polarization. In vivo and in vitro, most neurons develop a number of neurites from which one will be come the axon and the other dendrites. Only recently, the molecular mechanisms regulating neuronal polarity have begun to be described (Wiggin et al., 2005).
More than 20 years ago, Gary Banker pioneered the use of cultured hippocampal neurons as a model system to study axon–dendrite specification. This model permitted the discovery that neurons in vitro initially grow a number of similar neurites (stage 2) (Fig. 1A), and, after 24 h, one of the neurites begins to elongate very rapidly, becoming the axon and leading to the morphological polarization of the neuron (stage 3) (Fig. 1A). Because the neurite that shows the greatest growth will invariably become an axon, it has been hypothesized that axon specification and axon growth are governed in part by common pathways (Jiang and Rao, 2005). Earlier this year, Gary Banker's laboratory provided evidence that a kinesin-1 family member is targeted to the neurite that will start to elongate and become the axon (Jacobson et al., 2006). This work led the authors to propose that selective transport of kinesin-1 cargoes to the tip of a single neurite may contribute to axon specification. In a paper by the same laboratory in The Journal of Neuroscience, Oliva et al. (2006) go one step further by demonstrating that the kinase JNK (c-Jun N-terminal protein kinase), which is transported by a kinesin-1 cargo, is decisive in the regulation of axonal growth and specification during stage 2–3 transition.

{kind=link}
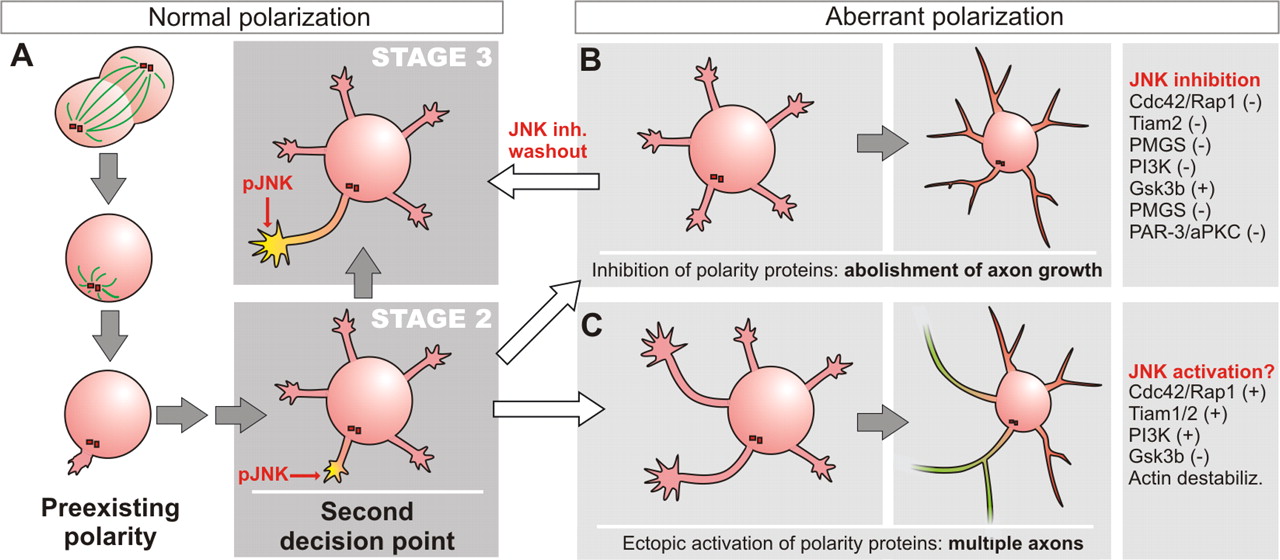
Stages of normal (A) and aberrant (B, C) hippocampal neuron differentiation in vitro. A, The cleavage plane of the last mitotic division determines the position of the centrosome, which primes the area of formation of the initial neurite. Shortly afterward, a number of similar neurites develop (stage 2). Although at this stage every neurite has the potential to become the axon, some proteins involved in axon development, such as active JNK, are already present in the initial neurite. In normal conditions, this is the neurite that will begin to elongate rapidly and form the axon (A). B, When some of the proteins required for axon formation are inhibited, no axon is formed and all of the neurites become dendrites. In the case of JNK, this aberrant polarization can be reverted if JNK inhibitor is removed from the medium. C, When the activity of these proteins is induced ectopically, axons develop from nonprimed neurites. Some examples of experiments that lead to aberrant polarization are shown at the right (for review, see Wiggin et al., 2005). Green neurite, Axon; red neurite, dendrite; −, inhibition; +, activation
Using the same model system, Oliva et al. investigated a number of protein kinases that might be involved in axon formation. The authors found that active, phosphorylated JNK was enriched in the axons of polarized neurons (Fig. 1A) [Oliva et al. (2006), their Fig. 1a,b (http://www.jneurosci.org/cgi/content/full/26/37/9462/F1)], although total JNK was distributed throughout the cell [Oliva et al. (2006), their Fig. 1d,e (http://www.jneurosci.org/cgi/content/full/26/37/9462/F1)]. Furthermore, they found that phosphorylated JNK was absent from neurites of most stage 2 neurons, before axon emergence, but became enriched in the nascent axon during stage 2–3 transition (Fig. 1A) [Oliva et al. (2006), their Fig. 1f,g (http://www.jneurosci.org/cgi/content/full/26/37/9462/F1)]. This selective activation of JNK in the growing axon suggested the participation of JNK in axon growth or specification. Indeed, Oliva et al. demonstrated, using pharmacological and dominant-negative approaches, that JNK activity is necessary for proper axon development. Pharmacological inhibition of JNK before neurite formation abolished neuronal polarity without preventing the formation of minor neurites (Fig. 1A) [Oliva et al. (2006), their Fig. 2 (http://www.jneurosci.org/cgi/content/full/26/37/9462/F2)], which would mature to become dendrites, displaying dendritic branching pattern and markers such as microtubule-associated protein MAP2 but lacking the axonal marker Tau-1 [Oliva et al. (2006), their Fig. 3 (http://www.jneurosci.org/cgi/content/full/26/37/9462/F3)]. Similar results were obtained using the JNK-binding domain of JIP-1 (JNK interacting protein 1) that acts as an inhibitor of JNK [Oliva et al. (2006), their Fig. 7 (http://www.jneurosci.org/cgi/content/full/26/37/9462/F7)], demonstrating that JNK activity is required for axon formation but not for neurite growth. Interestingly, many of the neurons that were grown during 6 d in the presence of JNK inhibitor and had formed dendrites (based on morphology and molecular markers) retained the capacity to develop a single axon 24 h after inhibitor removal (Fig. 1A) [Oliva et al. (2006), their Fig. 5 (http://www.jneurosci.org/cgi/content/full/26/37/9462/F5)]. Therefore, JNK may actually regulate axon formation by controlling axon growth and the expression of axonal markers but not axon fate, although these events are difficult to dissociate (Jiang and Rao, 2005).
How JNK activation regulates neuronal polarity is not clear. One possible effector is the activating transcription factor ATF-2, which is enriched in the growing axon and phosphorylated by JNK, as determined by pharmacological inhibition of JNK in cultured neurons [Oliva et al. (2006), their supplemental Fig. 3 (http://www.jneurosci.org/content/vol26/issue37/images/data/9462/DC1/OlivaSupplFig3.gif) and Fig. 8 (http://www.jneurosci.org/cgi/content/full/26/37/9462/F8)]. Although ATF-2 may promote axon elongation, gene transcription is not required for axon specification, as discussed by the authors, which is consistent with the hypothesis that JNK could become activated after axon specification and then promote axon growth and maturation. To distinguish between these two possibilities, inducing the activation of JNK in one of the minor processes would be critical, although such an experiment could not be attained by the authors for logistical reasons. If ectopic activation of JNK leads to the formation of an axon from a minor neurite, this would lend additional support to a model of JNK regulating axon specification.
{kind=link}
The piece that Oliva et al. introduce in the puzzle of neuronal polarity is a very special one. JNK targets include several actin and microtubule-associated proteins, such as Doublecortin, whose phosphorylation by JNK is necessary for proper axon growth (Gdalyahu et al., 2004). Because kinesin-1 translocation seems to be one of the earliest steps during stage 2–3 transition (Jacobson et al., 2006) and it transports JNK targets and JNK activating kinases, selective transport may ensure the activation of JNK pathway only at the axon-to-be neurite tip and initiate the growth and molecular differentiation required to become the axon. Indeed, as shown by the authors, if JNK fails to be activated, axon formation is abolished. Therefore, the JNK pathway can be added to those believed to regulate axon specification or formation during stage 2–3 transition (Fig. 1) (Wiggin et al., 2005).
Why this critical transition point exists is not clear. It has been established that the position of the centrosome, determined by the last mitotic division, defines the localization of the first neurite to develop, which will become the axon (Fig. 1) (Calderon de Anda et al., 2005). The centrosome polarizes microtubule polymerization and membrane transport, and therefore the proximal neurite potentially responds better to extrinsic cues (and elongates faster). However, this preexisting polarity is reconsidered during stage 2, at least in vitro, when all of the neurites display similar length and the potential to become the axon. Therefore, if during the stage 2–3 transition the primed neurite fails to elongate faster, or a minor neurite is induced to elongate, the original axon–dendrite fate can be reassigned (Fig. 1B,C) (Wiggin et al., 2005). This second decision point would grant the neuron an opportunity to select again the neurite most fit to the existing conditions, frequently the first neurite, to ensure the successful formation of an axon. It will be important not only to identify the set of proteins that control this transition in vitro and their interactions but also to understand how these pathways regulate polarity in vivo, and whether this second decision point also occurs.
Footnotes
- Received September 26, 2006.
- Revision received September 29, 2006.
- Accepted September 29, 2006.
-
A.M.-L.M. was supported by a European Molecular Biology Organization Long Term Fellowship and a Michael Smith Foundation for Health Research Postdoctoral Fellowship.
-
Editor's Note: These short reviews of a recent paper in the Journal, written exclusively by graduate students or postdoctoral fellows, are intended to mimic the journal clubs that exist in your own departments or institutions. For more information on the format and purpose of the Journal Club, please see http://www.jneurosci.org/misc/ifa_features.shtml.
- Correspondence should be addressed to Dr. Ana Mingorance-Le Meur, Department of Cellular and Physiological Sciences, University of British Columbia, 2350 Health Sciences Mall, Vancouver, British Columbia, Canada V6T 1Z3. alemeur{at}interchange.ubc.ca
- Copyright © 2006 Society for Neuroscience 0270-6474/06/2612104-02$15.00/0