

Abstract
The AMPA receptor, ubiquitous in brain, is termed “ionotropic” because it gates an ion channel directly. We found that an AMPA receptor can also modulate a G-protein to gate an ion channel indirectly. Glutamate applied to a retinal ganglion cell briefly suppresses the inward current through a cGMP-gated channel. AMPA and kainate also suppress the current, an effect that is blocked both by their general antagonist CNQX and also by the relatively specific AMPA receptor antagonist GYKI-52466. Neither NMDA nor agonists of metabotropic glutamate receptors are effective. The AMPA-induced suppression of the cGMP-gated current is blocked when the patch pipette includes GDP-β-S, whereas the suppression is irreversible when the pipette contains GTP-γ-S. This suggests a G-protein mediator, and, consistent with this, pertussis toxin blocks the current suppression. Nitric oxide (NO) donors induce the current suppressed by AMPA, and phosphodiesterase inhibitors prevent the suppression. Apparently, the AMPA receptor can exhibit a “metabotropic” activity that allows it to antagonize excitation evoked by NO.
Glutamate, the major excitatory transmitter in brain, activates a class of receptors termed “ionotropic” because they directly gate ion channels (Nakanishi, 1992; Seeburg, 1993; Hille, 1994; Riedel, 1996; Pass, 1998). Recently, an ionotropic glutamate receptor of the subclass that binds AMPA was discovered in cortical homogenates to also have a “metabotropic” function: it activates a G-protein to suppress adenylyl cyclase (Wang et al., 1997). We wondered whether the AMPA receptor has other metabotropic functions and whether these might include indirect gating of ion channels. We thought to probe for a metabotropic effect in a system suited for subsequent investigation of its role in neural integration.
We chose the retinal ganglion cell because the effects of stimulating its dendritic AMPA receptors can be investigated in a slice preparation (Aizenman et al., 1988; Mittman et al., 1990; Cohen et al., 1994;Leinders-Zufall et al., 1994; Taylor et al., 1995; Zhang et al., 1995;Coleman and Miller, 1998; Matsui et al., 1998) and because the role in neural integration could then be investigated in the intact retinain vitro. Certain ganglion cells express a cGMP-gated channel that causes an inward current when nitric oxide (NO) stimulates guanylyl cyclase to raise [cGMP] (Ahmad et al., 1994). The natural source of NO is probably a class of amacrine cell that stains intensely for NADPH diaphorase (NO synthase) (Sandell, 1985; Sager, 1986). Reasoning that a current stimulated by one signal (NO) ought to be antagonized by another, we tested the AMPA receptor and discovered that in retinal ganglion cells it can activate a G-protein to suppress the cGMP-gated current.
MATERIALS AND METHODS
Preparation and recording. Slices from adult rat retina were cut at 200 μm (Werblin, 1978) and viewed on a Zeiss (Oberkochen, Germany) upright microscope with differential interference contrast optics (×40 water-immersion objective). Ganglion cells were identified in the slice by their position and size. Membrane currents were recorded in the whole-cell configuration (Hamill et al., 1981) using a patch-clamp amplifier (Axopatch 200A; Axon Instruments, Foster City, CA) linked to a computer. The voltage-clamp procedures were controlled by the pClamp software (Axon Instruments). Data were low-pass filtered (four-pole Bessel type) with a cutoff frequency of 5 kHz and then digitized at 10 kHz by an analog-to-digital interface. All experiments were performed at room temperature (23–25°C).
Solutions and drugs. The control Ringer’s solution contained (in mm): NaCl, 135; KCl, 5; CaCl2, 1; MgCl2, 1; HEPES, 10; and glucose, 10. The solution was adjusted with NaOH to pH 7.4 and bubbled with oxygen. CoCl2 (1 mm), picrotoxin (100 μm), and strychnine (1 μm) were also added to block synaptic transmission. The recording pipette contained (in mm): CsCl, 140; CaCl2, 1; EGTA or BAPTA, 5; HEPES, 10; and Mg-ATP, 2. The solution was adjusted with CsOH to pH 7.4. Pipette resistance was ∼7 MΩ.
Test substances were applied through the bath [8-bromo-cGMP, 8-bromo-cAMP, 8-p-chlorophenylthio-cGMP, CNQX, GYKI-52466, α-methyl-4-caboxyphenylglycine (MCPG), α-cyclopropyl-4-phosphonophenylglycine (CPPG), 1-methyl-3-isobutylxanthine (IBMX), zaprinast, methylene blue, or sodium nitroprusside], via pressure ejection for 1 sec from a “puffer” pipette (glutamate, AMPA, kainate, NMDA,l-2-amino-4-phosphonobutyrate [l-AP-4], or 1S,3R-1-aminocyclopentane-trans-1,3-dicarboxylic acid [trans-(1S,3R)-ACPD]), or via the patch pipette (cGMP, GTP-γ-S, GDP-β-S, pertussis toxin, or cholera toxin). MCPG and CPPG were purchased from Tocris Cookson (Ballwin, MO). Other chemicals were from Sigma (St. Louis, MO).
RESULTS
Recording from ganglion cells in the slice preparation of adult rat retina, we confirmed the observations of Ahmad et al. (1994) that certain ganglion cells express a cGMP-gated current. When a membrane-permeant analog of cGMP (8-bromo-cGMP) was bath-applied at 1 mm to a ganglion cell voltage-clamped at −50 mV, there was a sustained inward current (217 ± 6 pA; mean ± SEM) (Fig.1a). This current disappeared in normal Ringer’s solution. The same current was obtained with another membrane-permeant analog of cGMP [1 mm8-p-chlorophenylthio-cGMP, which strongly resists hydrolysis by phosphodiesterase (PDE)] in the bath and also when the recording pipette contained cGMP (Fig. 1d). A permeant analog of cAMP was ineffective (n = 4). The cGMP-gated current was observed in approximately half of the cells (37 of 73).
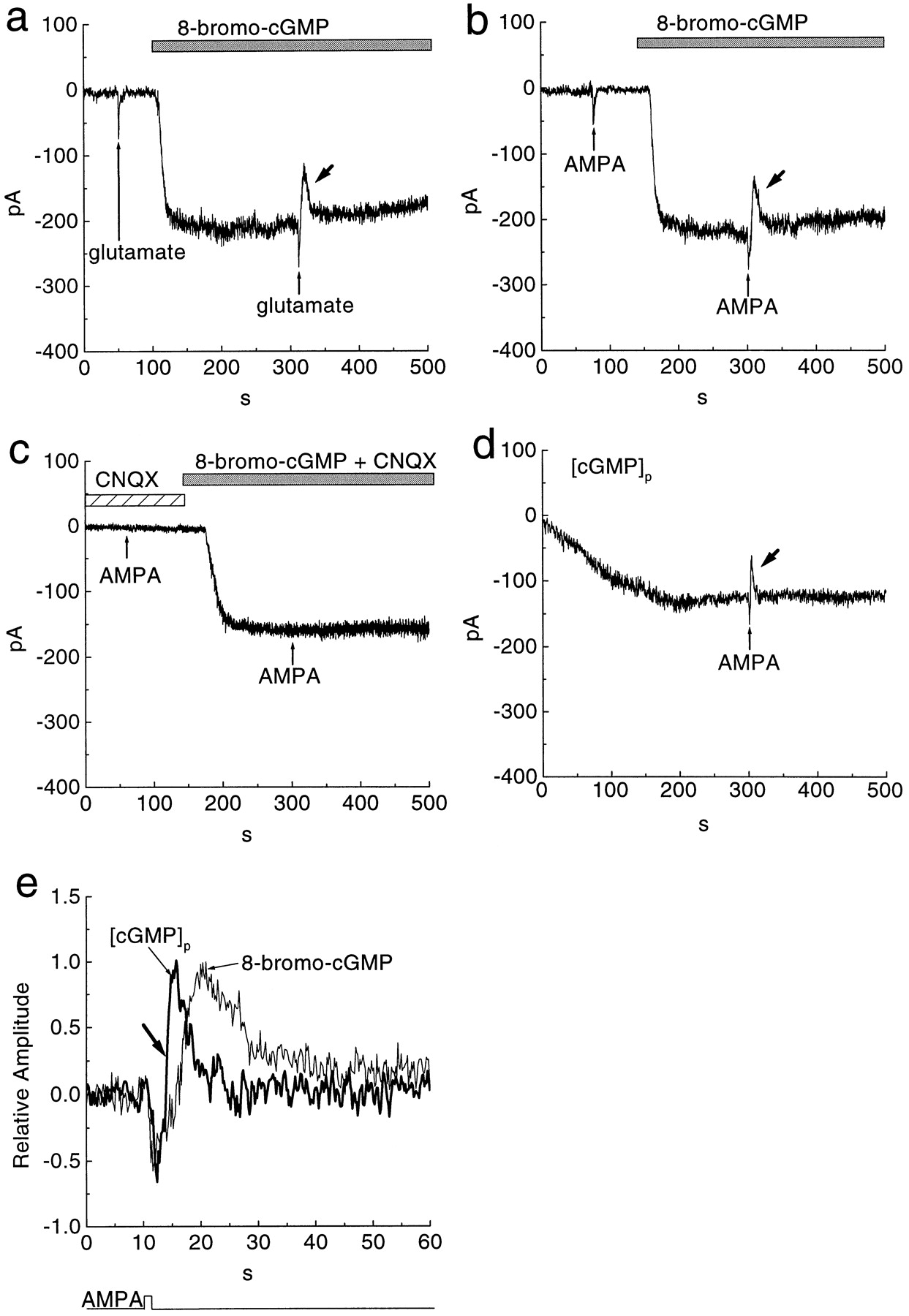
Stimulation of an AMPA receptor reduced a sustained inward current caused by cGMP. a,b, Puffer application of 100 μm glutamate for 1 sec (thin arrows) or 100 μm AMPA induced a fast transient inward current. Superfusion with 1 mm 8-bromo-cGMP (bars) activated a sustained inward current that was reduced (thick arrows) by puffer application of glutamate or AMPA. c, Application of 100 μm AMPA failed to reduce the 8-bromo-cGMP-induced sustained current in the solution containing 100 μm CNQX.d, Intracellular dialysis with 1 mm cGMP caused a slowly developing inward current that was also reduced (thick arrow) by puffer application of 100 μm AMPA. Whole-cell recording established att = 0. e, Normalized responses to AMPA application plotted from b and d on a faster scale. All cells were held at −50 mV.
When the sustained cGMP-gated current was induced by 8-bromo-cGMP, a puff of 100 μm glutamate evoked a biphasic response (Fig.1a). First, there was a transient inward current, as expected for direct gating of an ionotropic receptor; then, there was a brief reduction of the sustained inward current (41 ± 7%;n = 5) (Fig. 1a). The same biphasic response was evoked by AMPA (10–100 μm) (Fig. 1b) and by the AMPA receptor agonist kainate (10 μm;n = 3). The amplitude of the second phase (reduced inward current) was similar for 100 μm AMPA and 10 μm kainate (38 ± 6 and 34 ± 11%, respectively, with 8-bromo-cGMP). The amplitude of the second phase was also similar for 100 μm by AMPA and 100 μmglutamate (54 ± 7 and 55 ± 8%, respectively, with cGMP). However, when the sustained inward current was induced by 8-p-chlorophenylthio-cGMP, 100 μm AMPA did not reduce the second phase of the response (3 ± 1%;n = 3). The reduction by AMPA of the inward current was faster (Fig. 1e, thick arrow) when the sustained current was induced by cGMP rather than by 8-bromo-cGMP. The latency to maximal reduction after the AMPA puff was 4 ± 2 sec for cGMP and 11 ± 3 sec for 8-bromo-cGMP.
To identify which type of glutamate receptor reduced the inward current, we applied various agonists and antagonists of the ionotropic receptors. CNQX (100 μm), an antagonist of both AMPA and kainate subtypes, diminished the effect of AMPA (n = 4) (Fig. 1c). Furthermore, GYKI-52466 (100 μm), a specific antagonist of the AMPA receptor (Donevan and Rogawski, 1993), also diminished the effect of kainate (n = 3). Therefore, kainate receptors are probably not involved. The AMPA effects observed here, both the conventional transient inward current and the novel reduction of the sustained inward current, seem to desensitize rather little, as reported by others for AMPA responses of bipolar and ganglion cells in mammalian retina (Cohen et al., 1994;Sasaki and Kaneko, 1996). Finally, NMDA, applied as a 100 μm puff, evoked a monophasic inward current but did not reduce the inward current (n = 4) (Fig.2a). Thus, whatever causes this brief reduction of the inward current, it is apparently triggered specifically by the AMPA receptor.

The sustained inward current caused by cGMP was unaffected by NMDA and mGluR agonists. a, Application of 100 μm NMDA for 1 sec from a puffer pipette (arrows) failed to reduce the 8-bromo-cGMP-induced sustained current. b, Application of 100 μml-AP-4 (arrows) had no effect on a cell before or during superfusion of 1 mm8-bromo-cGMP. All cells were held at −50 mV.
To test whether a metabotropic glutamate receptor (mGluR) might reduce the inward current, we applied mGluR agonists. These includedl-AP-4 (n = 4) (Fig. 2b) and ACPD (n = 3). Although these compounds evoke large currents in ON bipolar cells (Nawy and Jahr, 1990; Shiells and Falk, 1990; Yamashita and Wässle, 1991; Tian and Slaughter, 1994) at the concentration used (100 μm), they did not affect the inward cGMP-gated current. Furthermore, mGluR antagonists MCPG and CPPG, both applied at 300 μm, did not affect the AMPA-evoked reduction of the steady inward current (n = 4). We conclude that the reduction by AMPA of the sustained inward current is not caused by activation of an mGluR.
If the reduction by AMPA of the sustained inward current results from closing the cGMP-gated channel, both the reduction and the sustained current should have the same reversal potential. The reduction by AMPA of the sustained inward current was outward at negative potentials and reversed at 0 mV (Fig. 3b). Similarly, the sustained current induced by 8-bromo-cGMP was inward at negative potentials and also reversed at approximately 0 mV (Fig.3b). Similar values were obtained from four cells (3 ± 4 mV), consistent with a previous report (Ahmad et al., 1994). Thus, both the cGMP-gated current and its AMPA-induced reduction reversed at the same membrane potential (n = 4) (Fig.3c), suggesting that AMPA suppresses the cGMP-gated current. If so, AMPA should also suppress the sustained cGMP-gated current induced by the NO donor, sodium nitroprusside, and we confirmed this (n = 4) (Fig. 4).

AMPA-induced reduction of current reversed at the same potential as the cGMP-gated current. a, Brief current evoked by a puff of 100 μm AMPA was inward at negative potentials and reversed at 0 mV. b, Same cell in Ringer’s solution containing 1 mm 8-bromo-cGMP. Slow transient current evoked by AMPA was outward at negative potentials and reversed at 0 mV. c, I–V curves of the steady and reduced cGMP-gated currents. Both currents reversed at 2 mV. These experiments suggest that AMPA actually suppresses the cGMP-gated current.

Superfusion with 100 μm sodium nitroprusside (SNP) activated a sustained inward current reduced by puffer application of 100 μm AMPA for 1 sec (arrows). A cell was held at −50 mV.
To investigate how the AMPA receptor suppresses the cGMP-gated current, we considered a role for Ca2+ as a second messenger (Kaupp and Koch, 1992; Nakanishi, 1992; Hille, 1994; Koutalos and Yau, 1996). If Ca2+ entering through the AMPA receptor was involved, removing extracellular Ca2+ should abolish the suppression. However, with 5 mm EGTA or BAPTA in a recording pipette, AMPA puffed onto a ganglion cell in Ca2+-free Ringer’s solution evoked the usual suppression of the cGMP-gated current (n = 6) (Fig.5a). AMPA (100 μm) reduced the current by 41 ± 7%, which was similar to that in control Ringer’s solution (38 ± 6%). The latency (12 ± 4 sec) to maximal current reduction after the AMPA puff was also similar to the control (11 ± 3 sec). When all permeant cations were removed from the medium, 8-bromo-cGMP induced no inward current at negative potentials; however, it did induce a sustained outward current at positive potentials, and this current was suppressed by AMPA. Thus, neither Ca2+ nor Na+ influx is required for the suppression, and we sought another mechanism.
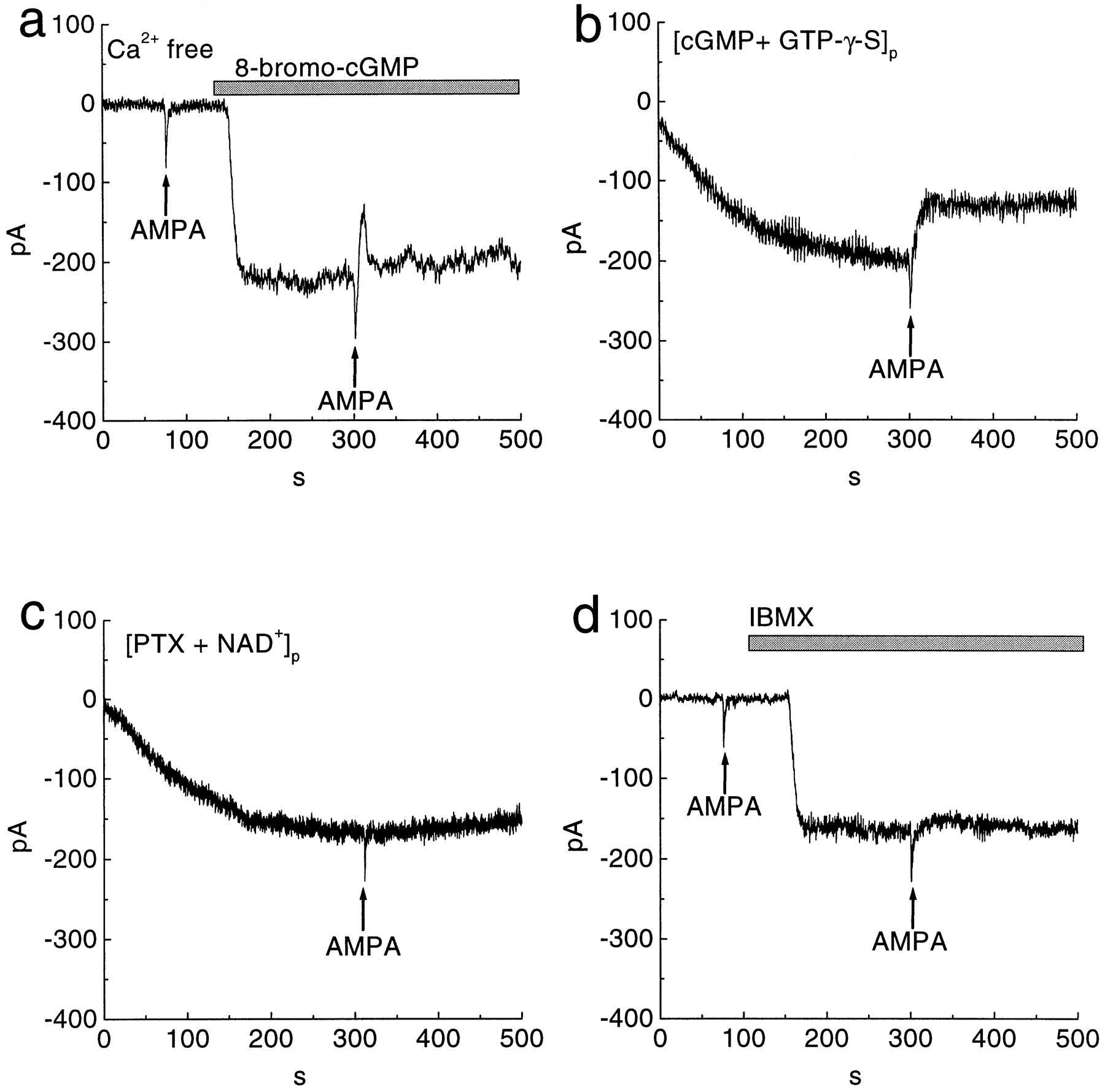
The suppression by AMPA of the cGMP-gated current is mediated by a G-protein that activates PDE. a, Application of 100 μm AMPA (arrows) suppressed the 8-bromo-cGMP-induced sustained current in the Ca2+-free solution. The recording pipette contained 5 mm EGTA. b, Dialysis of a cell with 1 mm cGMP and 100 μm GTP-γ-S produced an inward current. An AMPA puff (arrow) evoked a brief inward current and suppressed the sustained inward current irreversibly. Whole-cell recording established at t= 0. c, Dialysis of a cell with pertussis toxin (PTX; 1 μg/ml) and NAD+ (1 mm) also produced an inward current. An AMPA puff (arrow) evoked a brief inward current but failed to suppress the sustained inward current. d, Superfusion with 100 μm IBMX (bar) activated an inward current. An AMPA puff (arrows) also failed to suppress the inward current induced by IBMX. All cells were held at −50 mV.
We considered a role for a G-protein, possibly the Gifamily (Gi or Go), because AMPA can stimulate Gi (Wang et al., 1997) and because the inner plexiform and ganglion cell layers are immunoreactive for Gi/o (Terashima et al., 1987; Vardi et al., 1993; Oguni et al., 1996). To test for a G-protein activity, we used 1 mmcGMP in the patch pipette to cause a sustained inward current (as in Fig. 1d) but also included 100 μm GTP-γ-S. An AMPA puff induced the expected inward transient, followed by suppression of the sustained current. However, when the AMPA dissipated, the sustained current was not restored; rather, the suppression became essentially irreversible (n = 4) (Fig. 5b). This would be expected if an activated G-protein had bound GTP-γ-S because this GTP analog does not hydrolyze. In a separate experiment with 1 mm cGMP in the pipette, we used 500 μm GDP-β-S to competitively block G-protein activation (Nawy and Jahr, 1990). This produced a sustained inward current, which AMPA failed to suppress (n = 3).
Evidence that the G-protein species might be Gi/o came from experiments with pertussis toxin. Pertussis toxin ADP ribosylates Gi/o (Milligan, 1988), thereby blocking its downstream effect. Dialyzing a cell with pertussis toxin and NAD+ (omitting cGMP) produced a sustained inward current, which AMPA also failed to suppress (n = 5) (Fig. 5c). When NAD+ was omitted, pertussis toxin alone failed to produce the effect (n = 3), suggesting that ADP-ribosylation by pertussis toxin of Gi/o induced the inward current. This effect of pertussis toxin is consistent with previous results with ON bipolar cell (Shiells and Falk, 1992). Cholera toxin, which ADP ribosylates Gs, caused no inward current until 8-bromo-cGMP was added, and then the usual suppression was observed (n = 3). Because the cGMP-gated current seems to be suppressed by activating Gi/o, we considered possible intermediate effectors.
PDE seemed a natural candidate, by analogy with photoreceptors and certain bipolar cells in which a G-protein activates PDE to suppress a cGMP-gated current (Yau and Baylor, 1989; Kaupp and Koch, 1992; Lamb and Pugh, 1992; Koutalos and Yau, 1996). Furthermore, as noted, suppression by AMPA of the inward current was faster when the sustained current was induced by cGMP rather than by 8-bromo-cGMP (Fig.1e). This would be expected from a PDE mechanism because the brominated analog hydrolyzes slowly (Wei et al., 1998). The amplitude of the current suppression was approximately similar for both compounds. Although this might seem contradictory, the sluggish hydrolysis of PDE of 8-bromo-cGMP may be counterbalanced by its sixfold higher affinity for 8-bromo versus native cGMP (Wei et al., 1998). These considerations led us to try inhibitors of PDE.
We found, as first reported by Ahmad et al. (1994), that the PDE inhibitor IBMX induced a sustained inward current in a ganglion cell that expresses the cGMP-gated channel (n = 4) (Fig.5d). This implies that the cell also expresses an active PDE that regulates intracellular cGMP. With IBMX in the bath, an AMPA puff failed to suppress the inward current (n = 4) (Fig.5d). We obtained the same result using a selective inhibitor of cGMP phosphodiesterase, zaprinast (n = 3). Presumably AMPA was ineffective because Gi/o could not activate PDE. The suppression by AMPA of the cGMP-gated current was observed when, to the bath containing 8-bromo-cGMP, we added 1 mm methylene blue, which completely inhibits guanylyl cyclase (n = 4) (Danziger et al., 1993; Fratelli et al., 1995; Stuart-Smith et al., 1998). This result suggests that the suppression does not involve modulation of the cyclase.
DISCUSSION
In ganglion cells that express the cGMP-gated channel, AMPA acts in two ways: (1) transiently opens a cation channel; and (2) activates a G-protein that, via PDE, reduces [cGMP] to suppress a cGMP-gated current (Fig. 6). This second effect of AMPA cannot be attributed to activation of a genuine mGluR receptor because agonists and antagonists of mGluRs were ineffective. We feel confident that a PDE is involved because the effect was abolished by inhibitors of PDE and also by the analog of cGMP, which strongly resists hydrolysis. In short, this AMPA receptor seems to exhibit both ionotropic and metabotropic activities. This conclusion fits biochemical evidence that an AMPA receptor in cortex can activate a G-protein to modulate cAMP (Wang et al., 1997) and that a kainate receptor in hippocampus can activate a G-protein to modulate yet another biochemical cascade (phospholipase C; Rodriguez-Moreno and Lerma, 1998). Thus, metabotropic effects of ionotropic glutamate receptors may prove to be both widespread and diverse.

Model explains how the AMPA receptor in retinal ganglion cells might suppress the cGMP-gated current. NO released from amacrine cells raises [cGMP] (Sagar, 1986), inducing the cGMP-gated current (Ahmad et al., 1994). Ganglion cells express Gi/o-coupled metabotropic receptors (shown by aquestion mark), such as GABAB (Zhang et al., 1997) and D2-dopamine receptor (Djamgoz and Wagner, 1992). Unidentified link is shown with a broken arrow.GLU, Glutamate; R, G-protein-coupled receptor; G, Gi or Goprotein.
Given that the suppression by AMPA of the cGMP-gated current acts via a non-NMDA ionotropic glutamate receptor, can the family be identified? Probably, it is the AMPA family (GluR1–4) because of the following: (1) ganglion cells express mRNA for these subunits most strongly (Hamassaki-Britto et al., 1993); (2) the concentrations of AMPA (10–100 μm) were insufficient to activate native kainate receptors (Lerma et al., 1993; Clarke et al., 1997; Huettner, 1997); and (3) the effect was blocked by GYKI-52466, which is relatively specific for AMPA receptors. Which particular AMPA subunits (GluR1–4) suppress the cGMP-gated current cannot be determined until their specific agonists and antagonists become available.
AMPA receptors on CNS neurons desensitize rapidly (Colquhoun et al., 1992; Raman and Trussell, 1992; Seeburg, 1993). Therefore, it surprised us that, as in previous reports on mammalian retina (Cohen et al., 1994; Sasaki and Kaneko, 1996), puff-applied AMPA causes little obvious desensitization. However, the time course of our puff application is slow compared with the rapid-flow method (Colquhoun et al., 1992; Raman and Trussell, 1992) used to study desensitization. If our ganglion cells desensitize in milliseconds, we would not see the effect. Another possibility is that ganglion cells might express an alternatively spliced “flip” module of the AMPA receptor, which desensitizes more slowly than a receptor containing both “flip” and “flop” modules (Sommer et al., 1990; Seeburg, 1993). Finally, a heteromer of AMPA subunits (GluR1–4) and kainate subunits (GluR5–7 and KA1/2) might resist desensitization.
How the AMPA receptor couples to the G-protein remains to be established. Seven transmembrane receptors, including the mGluRs, contain a cytoplasmic tail with a conserved amino acid sequence that binds the G-protein (Nakanishi, 1992; Hille, 1994; Riedel, 1996; Pass, 1998). However, the AMPA receptor is assembled from hetero-oligomers that lack such a sequence (Sommer et al., 1990; Nakanishi, 1992;Seeburg, 1993; Pass, 1998), so the AMPA receptor probably does not affect the G-protein directly. Because there are now three examples in which AMPA and kainate receptor families use a G-protein to trigger a biochemical cascade (adenylyl cyclase, Wang et al., 1997; phospholipase C, Rodriguez-Moreno and Lerma, 1998; PDE, present study), it will be important to identify the coupling mechanism.
Approximately half of the recorded ganglion cells expressed the cGMP-gated current, and all of these showed the AMPA-induced suppression. We identified ganglion cells in the slice by their location (ganglion cell layer) and large size (compared with displaced amacrine cells) but did not study their morphology. It will now be interesting to determine which type(s) of ganglion cell express this mechanism and to learn its role in visual processing. Possibly, NO amacrine cells increase the cGMP-gated inward current of a ganglion cell and thus enhance spiking. In a preliminary experiment, we indeed observed that 8-bromo-cGMP had this effect. Glutamate from bipolar cells can suppress the cGMP-gated current to briefly curtail these extra spikes. Thus, the metabotropic action of the AMPA receptor might contribute to suppress excitation from the NO amacrine cells. Naturally, there are other possibilities, but this one emphasizes what may be a general mechanism in many brain regions: that glutamate can antagonize a key excitatory effect of NO.
Footnotes
This work was supported by National Institutes of Health Grant EY00828. We thank N. Vardi, S. Zigmond, P. Liebman, D. Manning, S. Nawy, and A. Kaneko for comments; R. Smith, M. Freed, L. Haarsma, J. Demb, and J. Tanaka for technical advice; and S. Watanabe for technical advice on the slice preparation.
Correspondence should be addressed to Dr. Fusao Kawai, c/o Dr. Peter Sterling, 123 Anatomy/Chemistry Building, Department of Neuroscience, School of Medicine, University of Pennsylvania, Philadelphia, PA 19104-6058.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}